

Follistatin-like 1 (Fstl1) is a bone morphogenetic protein (BMP) 4 signaling antagonist in controlling mouse lung development
- PMID: 21482757
- PMCID: PMC3084141
- DOI: 10.1073/pnas.1007293108
Follistatin-like 1 (Fstl1) is a bone morphogenetic protein (BMP) 4 signaling antagonist in controlling mouse lung development
Abstract
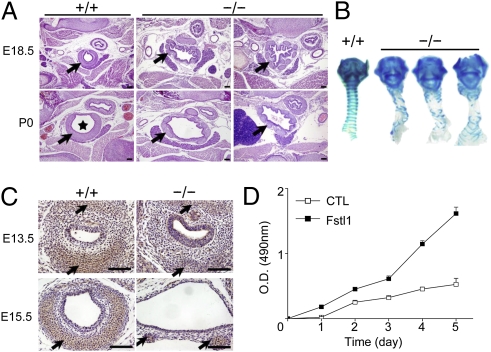
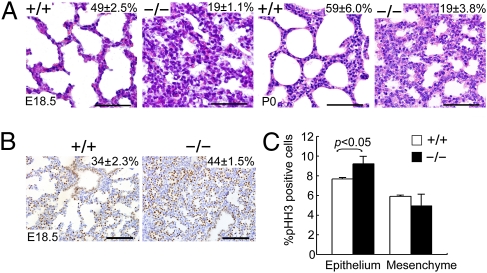
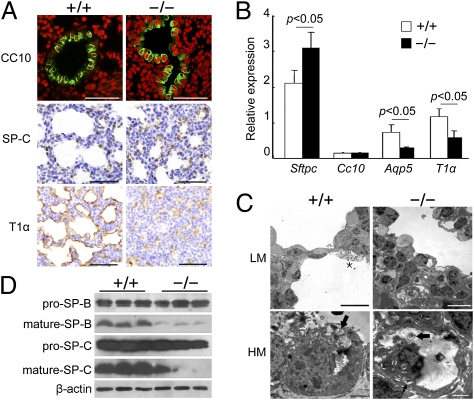
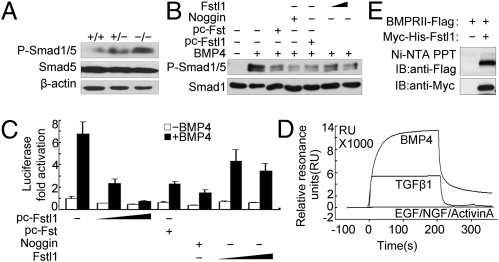
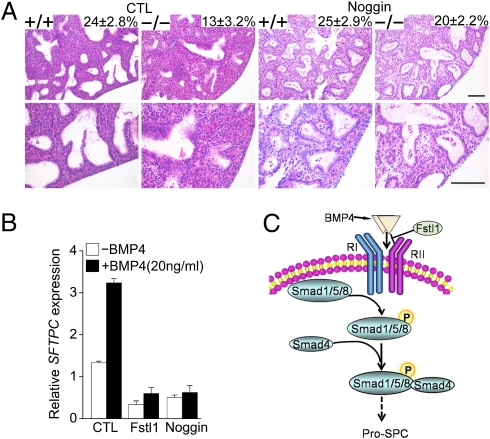
Lung morphogenesis is a well orchestrated, tightly regulated process through several molecular pathways, including TGF-β/bone morphogenetic protein (BMP) signaling. Alteration of these signaling pathways leads to lung malformation. We investigated the role of Follistatin-like 1 (Fstl1), a secreted follistatin-module-containing glycoprotein, in lung development. Deletion of Fstl1 in mice led to postnatal lethality as a result of respiratory failure. Analysis of the mutant phenotype showed that Fstl1 is essential for tracheal cartilage formation and alveolar maturation. Deletion of the Fstl1 gene resulted in malformed tracheal rings manifested as discontinued rings and reduced ring number. Fstl1-deficient mice displayed septal hypercellularity and end-expiratory atelectasis, which were associated with impaired differentiation of distal alveolar epithelial cells and insufficient production of mature surfactant proteins. Mechanistically, Fstl1 interacted directly with BMP4, negatively regulated BMP4/Smad1/5/8 signaling, and inhibited BMP4-induced surfactant gene expression. Reducing BMP signaling activity by Noggin rescued pulmonary atelectasis of Fstl1-deficient mice. Therefore, we provide in vivo and in vitro evidence to demonstrate that Fstl1 modulates lung development and alveolar maturation, in part, through BMP4 signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Maeda Y, Davé V, Whitsett JA. Transcriptional control of lung morphogenesis. Physiol Rev. 2007;87:219–244. - PubMed
-
- Rossant J, Tam PPL. Mouse Development: Patterning, Morphogenesis, and Organogenesis. Amsterdam: Elsevier; 2002.
-
- Que J, Choi M, Ziel JW, Klingensmith J, Hogan BL. Morphogenesis of the trachea and esophagus: Current players and new roles for noggin and Bmps. Differentiation. 2006;74:422–437. - PubMed
-
- Hogan BL, Yingling JM. Epithelial/mesenchymal interactions and branching morphogenesis of the lung. Curr Opin Genet Dev. 1998;8:481–486. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
