

Cargo binding activates myosin VIIA motor function in cells
- PMID: 21482763
- PMCID: PMC3084129
- DOI: 10.1073/pnas.1009188108
Cargo binding activates myosin VIIA motor function in cells
Abstract
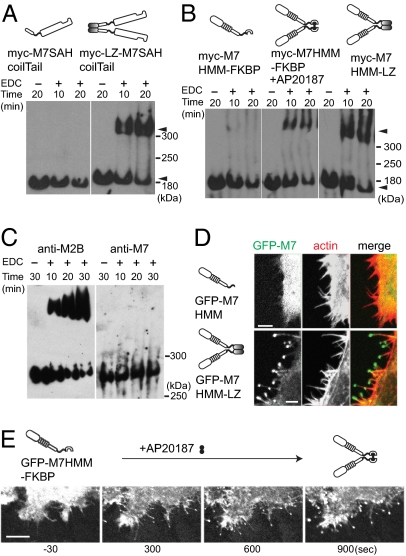
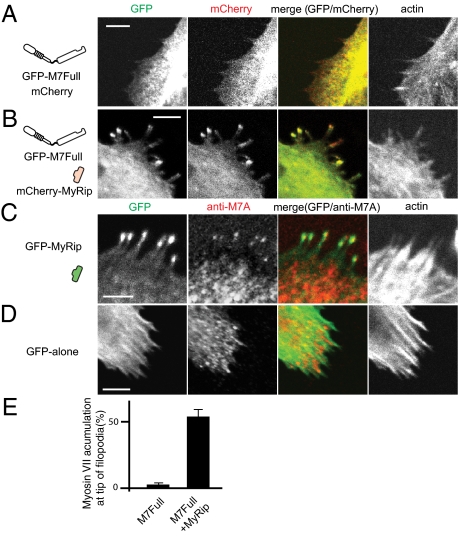
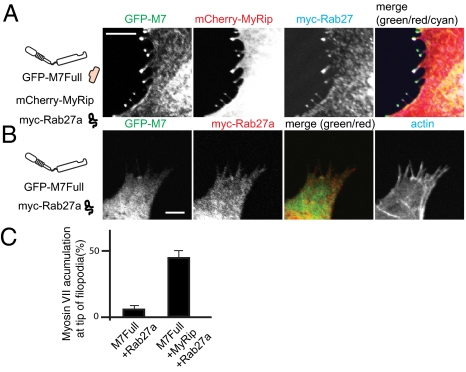
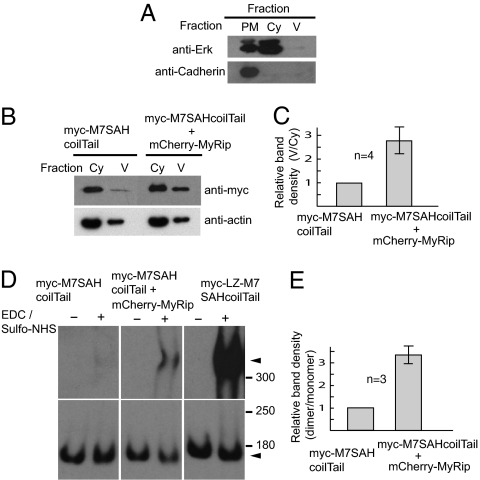
Myosin VIIA, thought to be involved in human auditory function, is a gene responsible for human Usher syndrome type 1B, which causes hearing and visual loss. Recent studies have suggested that it can move processively if it forms a dimer. Nevertheless, it exists as a monomer in vitro, unlike the well-known two-headed processive myosin Va. Here we studied the molecular mechanism, which is currently unknown, of activating myosin VIIA as a cargo-transporting motor. Human myosin VIIA was present throughout cytosol, but it moved to the tip of filopodia upon the formation of dimer induced by dimer-inducing reagent. The forced dimer of myosin VIIA translocated its cargo molecule, MyRip, to the tip of filopodia, whereas myosin VIIA without the forced dimer-forming module does not translocate to the filopodial tips. These results suggest that dimer formation of myosin VIIA is important for its cargo-transporting activity. On the other hand, myosin VIIA without the forced dimerization module became translocated to the filopodial tips in the presence of cargo complex, i.e., MyRip/Rab27a, and transported its cargo complex to the tip. Coexpression of MyRip promoted the association of myosin VIIA to vesicles and the dimer formation. These results suggest that association of myosin VIIA monomers with membrane via the MyRip/Rab27a complex facilitates the cargo-transporting activity of myosin VIIA, which is achieved by cluster formation on the membrane, where it possibly forms a dimer. Present findings support that MyRip, a cargo molecule, functions as an activator of myosin VIIA transporter function.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Weil D, et al. Defective myosin VIIA gene responsible for Usher syndrome type 1B. Nature. 1995;374:60–61. - PubMed
-
- Weil D, et al. The autosomal recessive isolated deafness, DFNB2, and the Usher 1B syndrome are allelic defects of the myosin-VIIA gene. Nat Genet. 1997;16:191–193. - PubMed
-
- Liu XZ, Newton VE, Steel KP, Brown SD. Identification of a new mutation of the myosin VII head region in Usher syndrome type 1. Hum Mutat. 1997;10:168–170. - PubMed
-
- Liu XZ, et al. Mutations in the myosin VIIA gene cause non-syndromic recessive deafness. Nat Genet. 1997;16:188–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
