

Design of a switchable eliminase
- PMID: 21482808
- PMCID: PMC3084051
- DOI: 10.1073/pnas.1018191108
Design of a switchable eliminase
Abstract
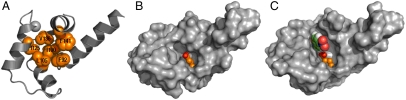
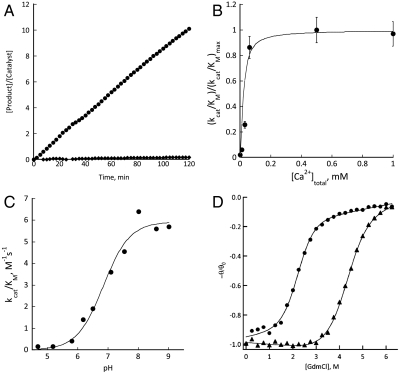
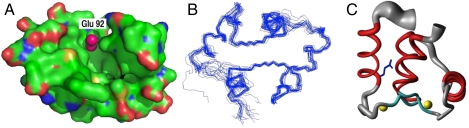
The active sites of enzymes are lined with side chains whose dynamic, geometric, and chemical properties have been finely tuned relative to the corresponding residues in water. For example, the carboxylates of glutamate and aspartate are weakly basic in water but become strongly basic when dehydrated in enzymatic sites. The dehydration of the carboxylate, although intrinsically thermodynamically unfavorable, is achieved by harnessing the free energy of folding and substrate binding to reach the required basicity. Allosterically regulated enzymes additionally rely on the free energy of ligand binding to stabilize the protein in a catalytically competent state. We demonstrate the interplay of protein folding energetics and functional group tuning to convert calmodulin (CaM), a regulatory binding protein, into AlleyCat, an allosterically controlled eliminase. Upon binding Ca(II), native CaM opens a hydrophobic pocket on each of its domains. We computationally identified a mutant that (i) accommodates carboxylate as a general base within these pockets, (ii) interacts productively in the Michaelis complex with the substrate, and (iii) stabilizes the transition state for the reaction. Remarkably, a single mutation of an apolar residue at the bottom of an otherwise hydrophobic cavity confers catalytic activity on calmodulin. AlleyCat showed the expected pH-rate profile, and it was inactivated by mutation of its active site Glu to Gln. A variety of control mutants demonstrated the specificity of the design. The activity of this minimal 75-residue allosterically regulated catalyst is similar to that obtained using more elaborate computational approaches to redesign complex enzymes to catalyze the Kemp elimination reaction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Broo KS, Brive L, Ahlberg P, Baltzer L. Catalysis of hydrolysis and transesterification reactions of p-Nitrophenyl esters by a designed helix—loop—helix dimer. J Am Chem Soc. 1997;119:11362–11372.
-
- Johnsson K, Allemann RK, Widmer H, Benner SA. Synthesis, structure and activity of artificial, rationally designed catalytic polypeptides. Nature. 1993;365:530–532. - PubMed
-
- Casey M, Kemp D, Paul K, Cox D. Physical organic chemistry of benzisoxazoles. I. Mechanism of the base-catalyzed decomposition of benzisoxazoles. J Org Chem. 1973;38:2294–2301.
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
