

The Dsl1 tethering complex actively participates in soluble NSF (N-ethylmaleimide-sensitive factor) attachment protein receptor (SNARE) complex assembly at the endoplasmic reticulum in Saccharomyces cerevisiae
- PMID: 21482823
- PMCID: PMC3137076
- DOI: 10.1074/jbc.M110.215657
The Dsl1 tethering complex actively participates in soluble NSF (N-ethylmaleimide-sensitive factor) attachment protein receptor (SNARE) complex assembly at the endoplasmic reticulum in Saccharomyces cerevisiae
Abstract
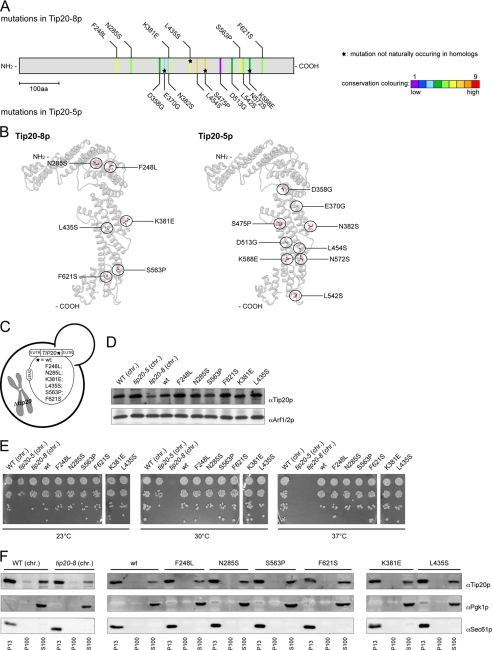
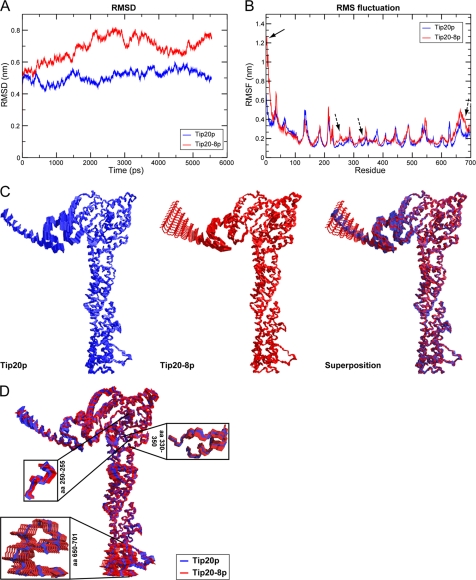
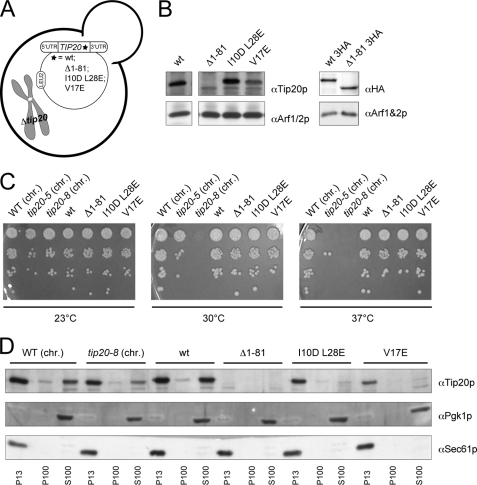
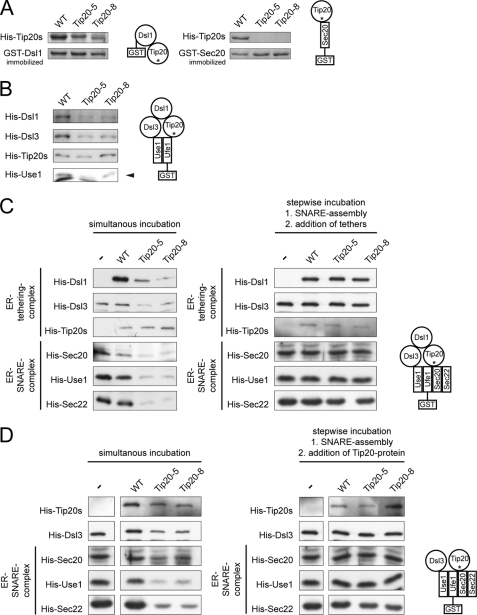
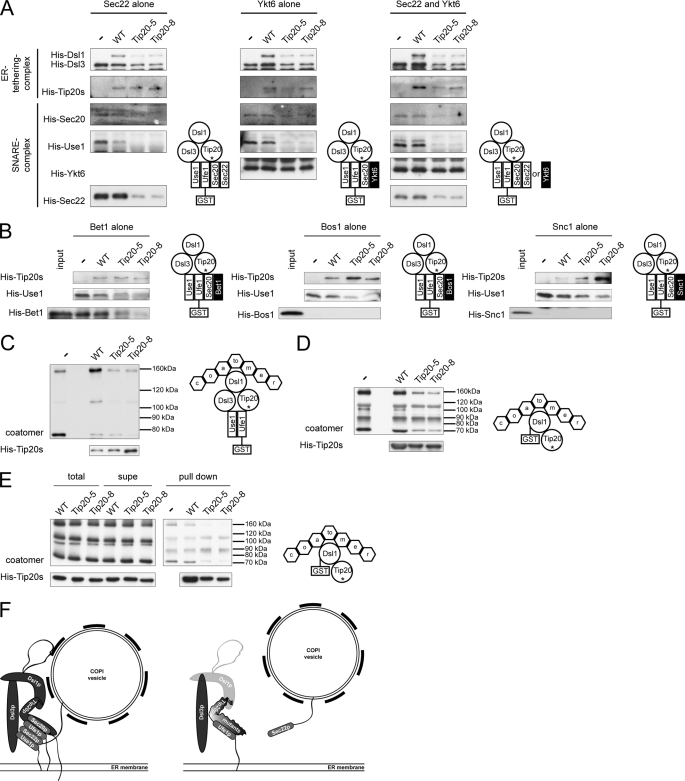
Intracellular transport is largely dependent on vesicles that bud off from one compartment and fuse with the target compartment. The first contact of an incoming vesicle with the target membrane is mediated by tethering factors. The tethering factor responsible for recruiting Golgi-derived vesicles to the ER is the Dsl1 tethering complex, which is comprised of the essential proteins Dsl1p, Dsl3p, and Tip20p. We investigated the role of the Tip20p subunit at the ER by analyzing two mutants, tip20-5 and tip20-8. Both mutants contained multiple mutations that were scattered throughout the TIP20 sequence. Individual mutations could not reproduce the temperature-sensitive phenotype of tip20-5 and tip20-8, indicating that the overall structure of Tip20p might be altered in the mutants. Using molecular dynamics simulations comparing Tip20p and Tip20-8p revealed that some regions, particularly the N-terminal domain and parts of the stalk region, were more flexible in the mutant protein, consistent with its increased susceptibility to proteolysis. Both Tip20-5p and Tip20-8p mutants prevented proper ER trans-SNARE complex assembly in vitro. Moreover, Tip20p mutant proteins disturbed the interaction between Dsl1p and the coatomer coat complex, indicating that the Dsl1p-coatomer interaction could be stabilized or regulated by Tip20p. We provide evidence for a direct role of the Dsl1 complex, in particular Tip20p, in the formation and stabilization of ER SNARE complexes.
Figures
Similar articles
-
The Dsl1 protein tethering complex is a resident endoplasmic reticulum complex, which interacts with five soluble NSF (N-ethylmaleimide-sensitive factor) attachment protein receptors (SNAREs): implications for fusion and fusion regulation.J Biol Chem. 2011 Jul 15;286(28):25039-46. doi: 10.1074/jbc.M110.215327. Epub 2011 May 6. J Biol Chem. 2011. PMID: 21550981 Free PMC article.
-
The coatomer-interacting protein Dsl1p is required for Golgi-to-endoplasmic reticulum retrieval in yeast.J Biol Chem. 2001 Oct 19;276(42):39150-60. doi: 10.1074/jbc.M105833200. Epub 2001 Aug 7. J Biol Chem. 2001. PMID: 11493604
-
The Saccharomyces cerevisiae early secretion mutant tip20 is synthetic lethal with mutants in yeast coatomer and the SNARE proteins Sec22p and Ufe1p.Yeast. 1998 May;14(7):633-46. doi: 10.1002/(SICI)1097-0061(199805)14:7<633::AID-YEA267>3.0.CO;2-3. Yeast. 1998. PMID: 9639310
-
Dsl1p/Zw10: common mechanisms behind tethering vesicles and microtubules.Trends Cell Biol. 2010 May;20(5):257-68. doi: 10.1016/j.tcb.2010.02.001. Epub 2010 Mar 11. Trends Cell Biol. 2010. PMID: 20226673 Review.
-
Stx5-Mediated ER-Golgi Transport in Mammals and Yeast.Cells. 2019 Jul 26;8(8):780. doi: 10.3390/cells8080780. Cells. 2019. PMID: 31357511 Free PMC article. Review.
Cited by
-
ER arrival sites for COPI vesicles localize to hotspots of membrane trafficking.EMBO J. 2016 Sep 1;35(17):1935-55. doi: 10.15252/embj.201592873. Epub 2016 Jul 20. EMBO J. 2016. PMID: 27440402 Free PMC article.
-
The Exocyst Subunit Sec6 Interacts with Assembled Exocytic SNARE Complexes.J Biol Chem. 2015 Nov 20;290(47):28245-28256. doi: 10.1074/jbc.M115.673806. Epub 2015 Oct 7. J Biol Chem. 2015. PMID: 26446795 Free PMC article.
-
ER-associated retrograde SNAREs and the Dsl1 complex mediate an alternative, Sey1p-independent homotypic ER fusion pathway.Mol Biol Cell. 2014 Nov 1;25(21):3401-12. doi: 10.1091/mbc.E14-07-1220. Epub 2014 Sep 3. Mol Biol Cell. 2014. PMID: 25187651 Free PMC article.
-
The Comparative Genomics of Botryosphaeriaceae Suggests Gene Families of Botryosphaeria dothidea Related to Pathogenicity on Chinese Hickory Tree.J Fungi (Basel). 2024 Apr 22;10(4):299. doi: 10.3390/jof10040299. J Fungi (Basel). 2024. PMID: 38667970 Free PMC article.
-
Retrograde traffic from the Golgi to the endoplasmic reticulum.Cold Spring Harb Perspect Biol. 2013 Jun 1;5(6):a013391. doi: 10.1101/cshperspect.a013391. Cold Spring Harb Perspect Biol. 2013. PMID: 23732476 Free PMC article. Review.
References
-
- Bonifacino J. S., Glick B. S. (2004) Cell 116, 153–166 - PubMed
-
- Lee M. C., Miller E. A., Goldberg J., Orci L., Schekman R. (2004) Annu. Rev. Cell Dev. Biol. 20, 87–123 - PubMed
-
- Springer S., Spang A., Schekman R. (1999) Cell 97, 145–148 - PubMed
-
- Barlowe C., Orci L., Yeung T., Hosobuchi M., Hamamoto S., Salama N., Rexach M. F., Ravazzola M., Amherdt M., Schekman R. (1994) Cell 77, 895–907 - PubMed
-
- Letourneur F., Gaynor E. C., Hennecke S., Démollière C., Duden R., Emr S. D., Riezman H., Cosson P. (1994) Cell 79, 1199–1207 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
