

Development of extraocular muscles requires early signals from periocular neural crest and the developing eye
- PMID: 21482859
- PMCID: PMC3248700
- DOI: 10.1001/archophthalmol.2011.75
Development of extraocular muscles requires early signals from periocular neural crest and the developing eye
Abstract
Objectives: To identify and explain morphologic changes of the extraocular muscles (EOMs) in anophthalmic patients.
Methods: Retrospective medical record review of patients with congenital anophthalmia, using magnetic resonance imaging and intraoperative findings to characterize EOM morphology. We then used molecular biology techniques in zebrafish and chick embryos to determine the relationships among the developing eye, periocular neural crest, and EOMs.
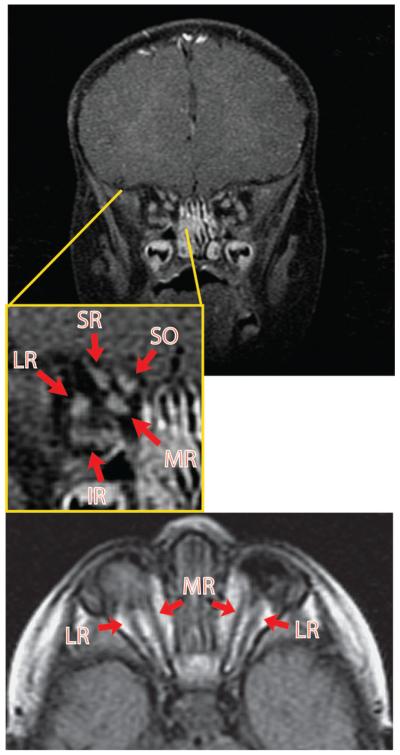
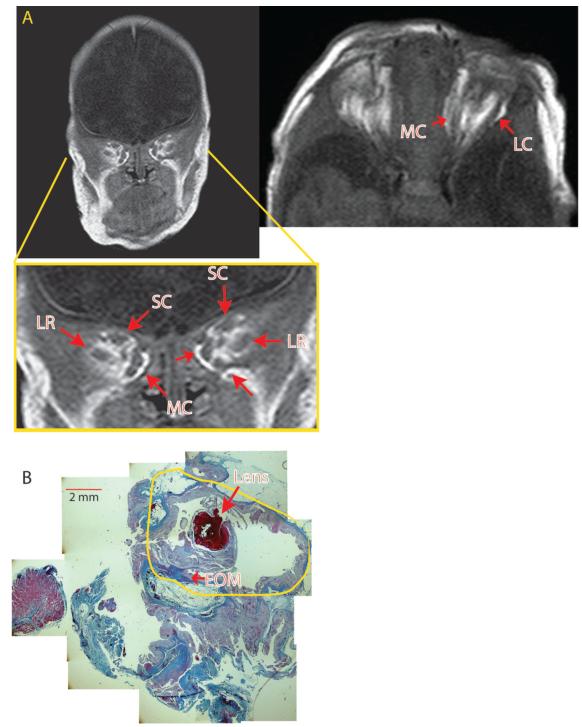
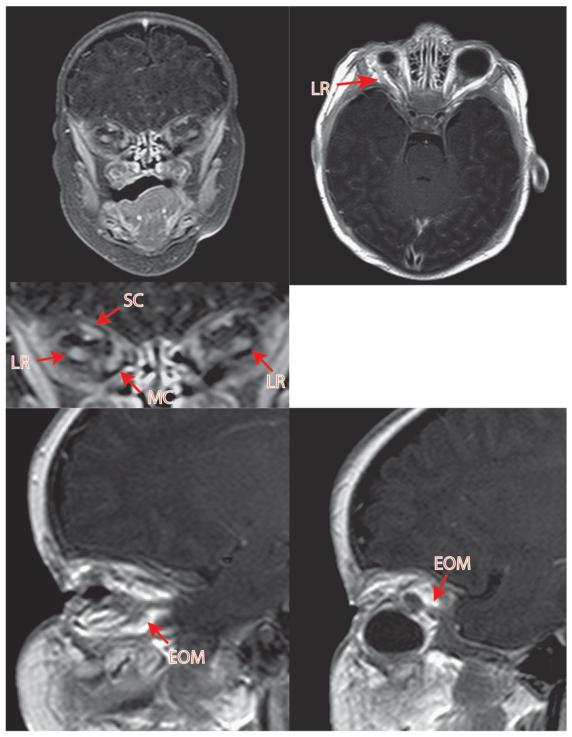
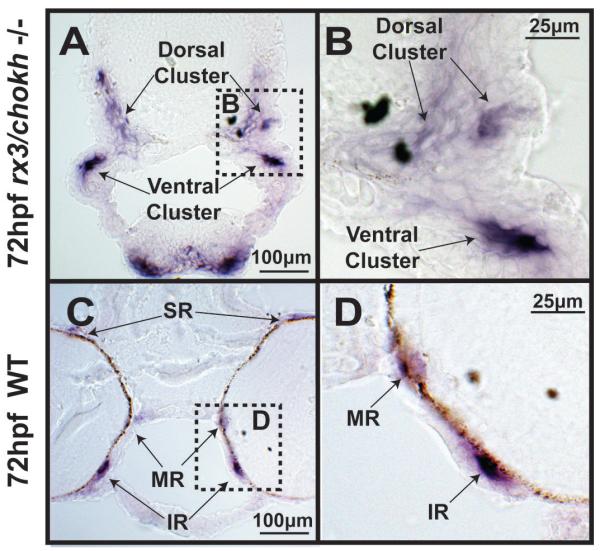
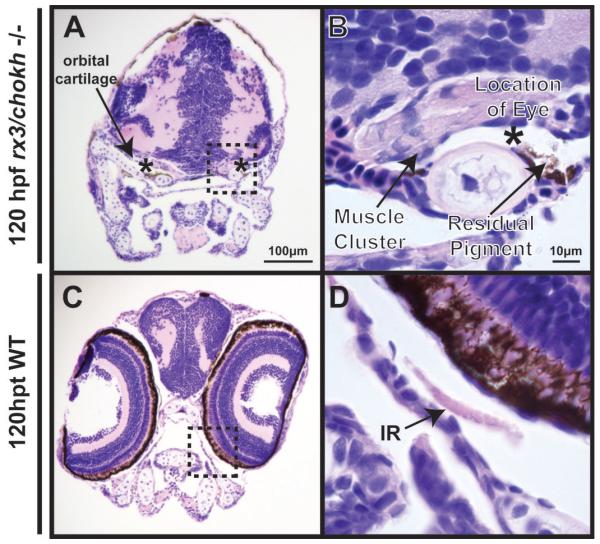
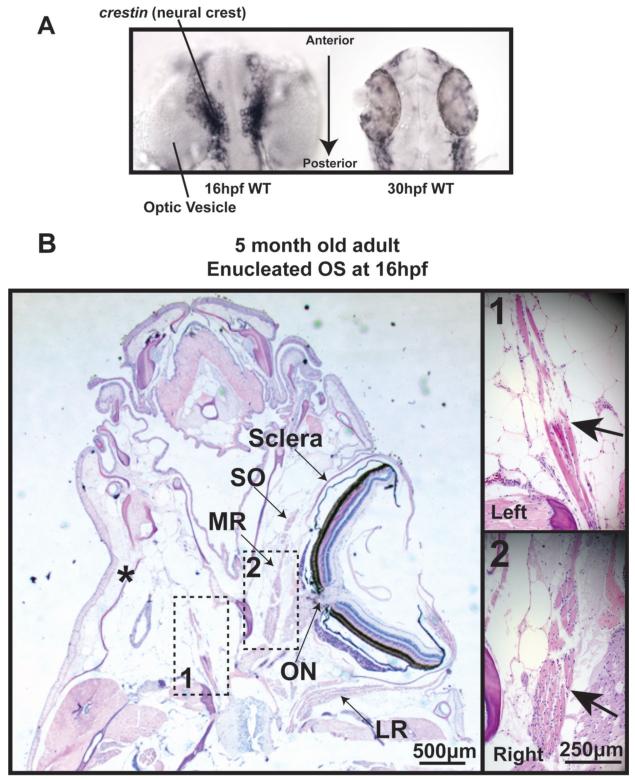
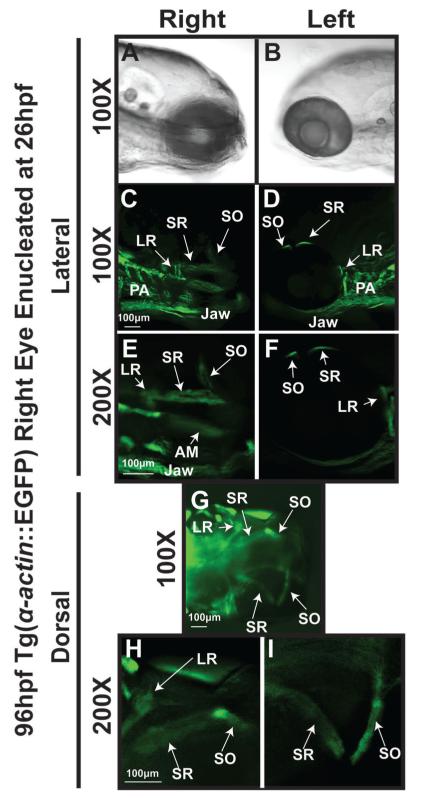
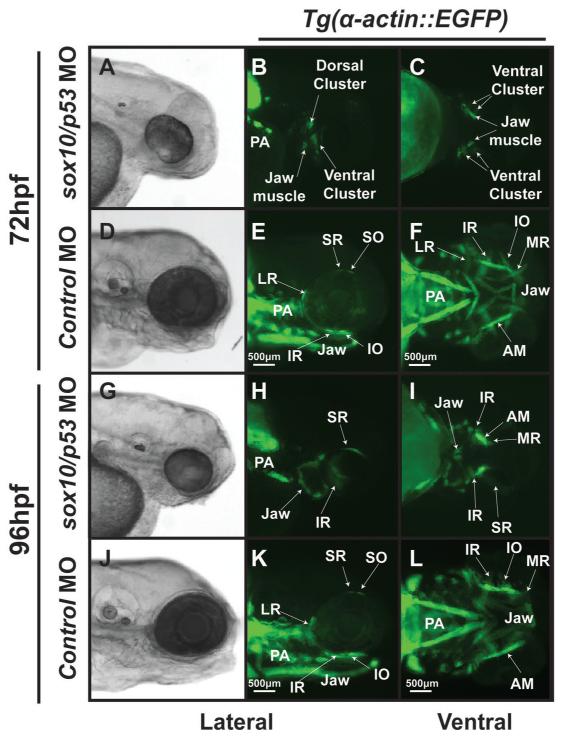
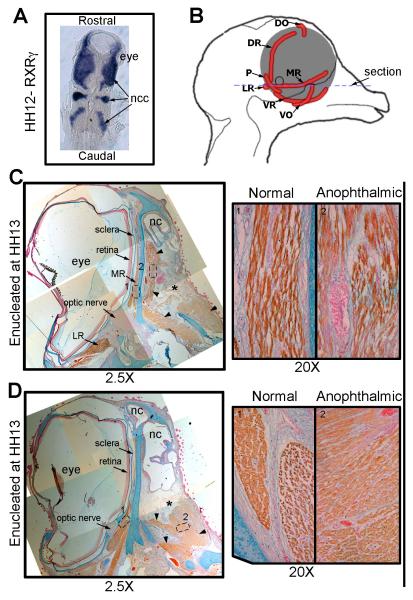
Results: In 3 human patients with bilateral congenital anophthalmia and preoperative orbital imaging, we observed a spectrum of EOM morphologies ranging from indiscernible muscle tissue to well-formed, organized EOMs. Timing of eye loss in zebrafish and chick embryos correlated with the morphology of EOM organization in the orbit (eye socket). In congenitally eyeless Rx3 zebrafish mutants, or following genetic ablation of the cranial neural crest cells, EOMs failed to organize, which was independent of other craniofacial muscle development.
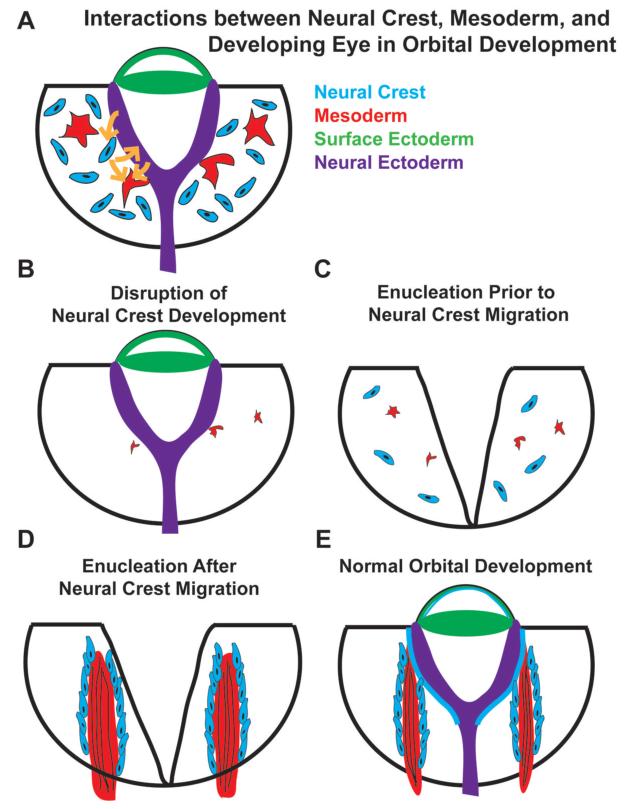
Conclusions: Orbital development is dependent on interactions between the eye, neural crest, and developing EOMs. Timing of the ocular insult in relation to neural crest migration and EOM development is a key determinant of aberrant EOM organization. Additional research will be required to study patients with unilateral and syndromic anophthalmia and assess for possible differences in clinical outcomes of patients with varied EOM morphology.
Clinical relevance: The presence and organization of EOMs in anophthalmic eye sockets may serve as a markers for the timing of genetic or teratogenic insults, improving genetic counseling, and assisting with surgical reconstruction and family counseling efforts.
Figures

Comment in
-
What experimental embryology can teach us about the development of the extraocular muscles in anophthalmia: at the interface of basic and clinical sciences.Arch Ophthalmol. 2011 Aug;129(8):1077-9. doi: 10.1001/archophthalmol.2011.187. Arch Ophthalmol. 2011. PMID: 21825193 Free PMC article. No abstract available.
References
-
- Porter JD, Israel S, Gong B, et al. Distinctive morphological and gene/protein expression signatures during myogenesis in novel cell lines from extraocular and hindlimb muscle. Physiol Genomics. 2006;24(3):264–275. - PubMed
-
- Yu Wai, Man CY, Chinnery PF, Griffiths PG. Extraocular muscles have fundamentally distinct properties that make them selectively vulnerable to certain disorders. Neuromuscul Disord. 2005;1(1):17–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
