

Spatial extent of charge repulsion regulates assembly pathways for lysozyme amyloid fibrils
- PMID: 21483680
- PMCID: PMC3071722
- DOI: 10.1371/journal.pone.0018171
Spatial extent of charge repulsion regulates assembly pathways for lysozyme amyloid fibrils
Abstract
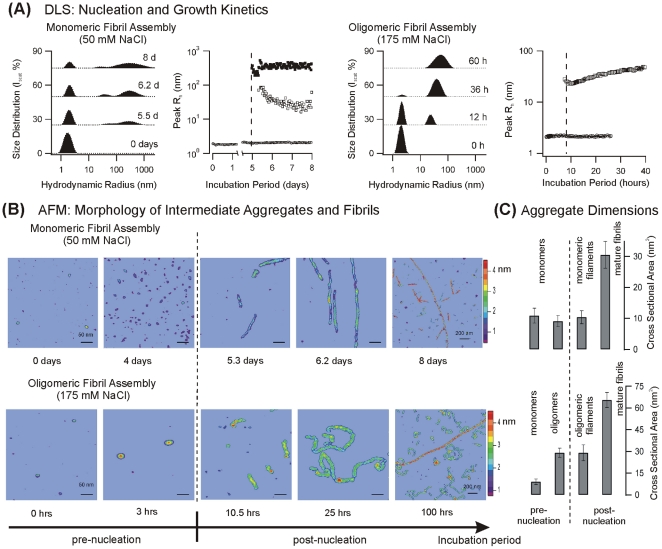
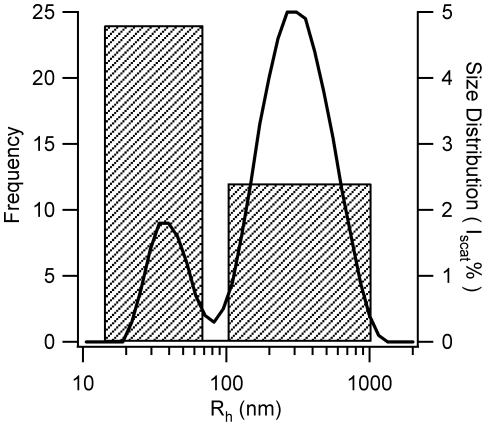
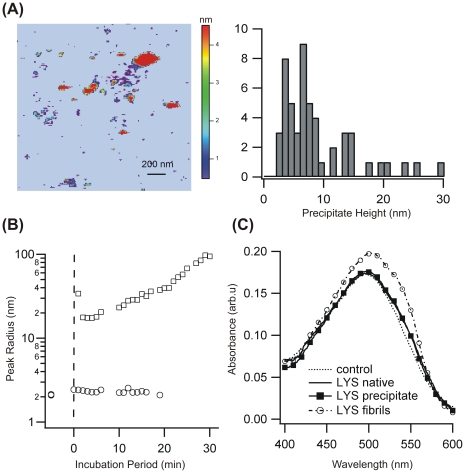
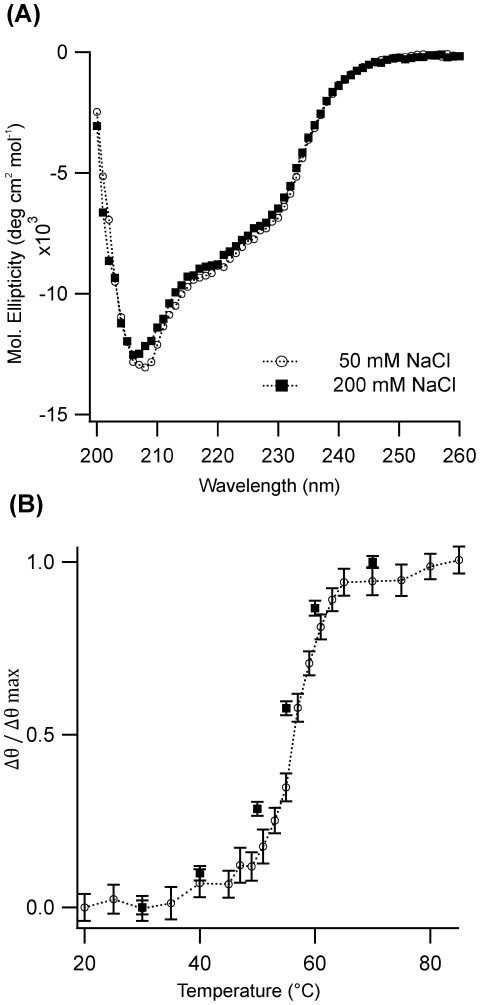
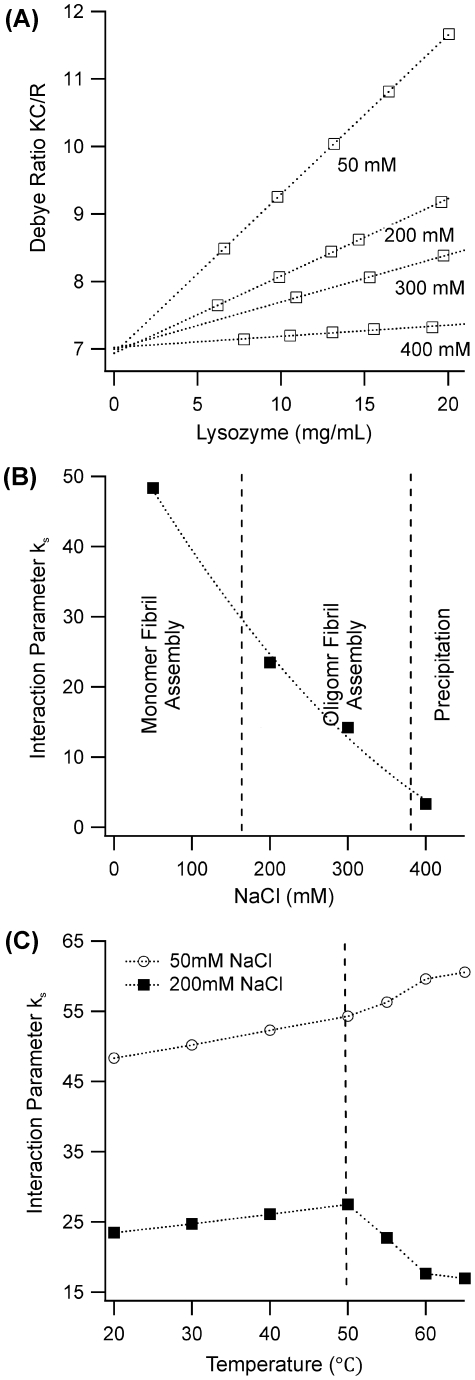
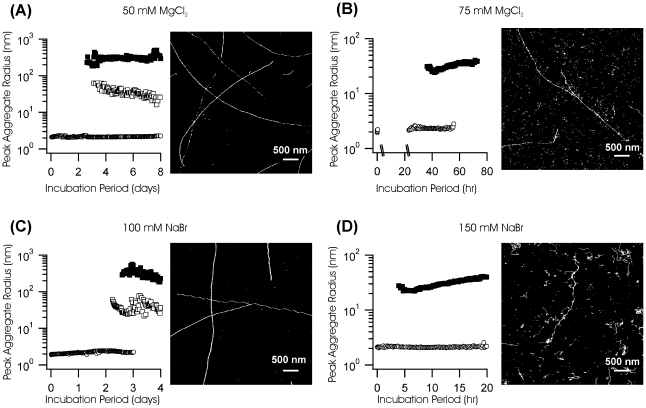
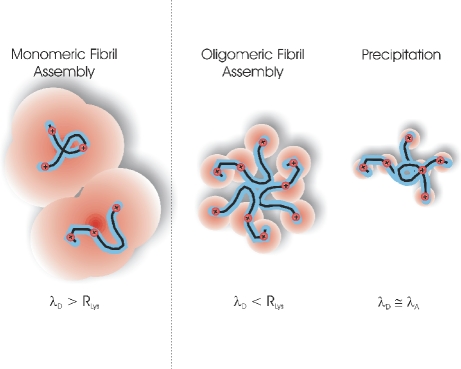
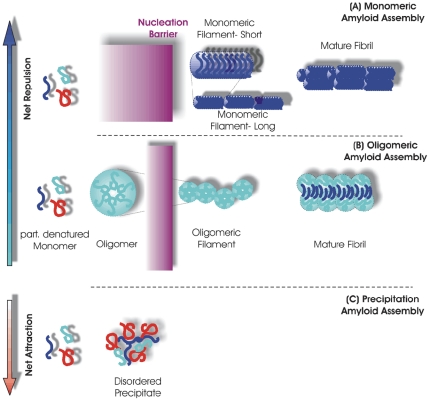
Formation of large protein fibrils with a characteristic cross β-sheet architecture is the key indicator for a wide variety of systemic and neurodegenerative amyloid diseases. Recent experiments have strongly implicated oligomeric intermediates, transiently formed during fibril assembly, as critical contributors to cellular toxicity in amyloid diseases. At the same time, amyloid fibril assembly can proceed along different assembly pathways that might or might not involve such oligomeric intermediates. Elucidating the mechanisms that determine whether fibril formation proceeds along non-oligomeric or oligomeric pathways, therefore, is important not just for understanding amyloid fibril assembly at the molecular level but also for developing new targets for intervening with fibril formation. We have investigated fibril formation by hen egg white lysozyme, an enzyme for which human variants underlie non-neuropathic amyloidosis. Using a combination of static and dynamic light scattering, atomic force microscopy and circular dichroism, we find that amyloidogenic lysozyme monomers switch between three different assembly pathways: from monomeric to oligomeric fibril assembly and, eventually, disordered precipitation as the ionic strength of the solution increases. Fibril assembly only occurred under conditions of net repulsion among the amyloidogenic monomers while net attraction caused precipitation. The transition from monomeric to oligomeric fibril assembly, in turn, occurred as salt-mediated charge screening reduced repulsion among individual charged residues on the same monomer. We suggest a model of amyloid fibril formation in which repulsive charge interactions are a prerequisite for ordered fibril assembly. Furthermore, the spatial extent of non-specific charge screening selects between monomeric and oligomeric assembly pathways by affecting which subset of denatured states can form suitable intermolecular bonds and by altering the energetic and entropic requirements for the initial intermediates emerging along the monomeric vs. oligomeric assembly path.
Conflict of interest statement
Figures
References
-
- Ross CA, Poirier MA. Protein aggregation and neurodegenerative disease. Nat Med. 2004;10:S10–S17. - PubMed
-
- Lansbury PT, Lashuel HA. A century-old debate on protein aggregation and neurodegeneration enters the clinic. Nature. 2006;443:774. - PubMed
-
- Chiti F, Dobson CM. Protein Misfolding, Functional Amyloid, and Human Disease. Annu Rev Biochem. 2006;75:333–366. - PubMed
-
- Kelly JW. Alternative conformations of amyloidogenic proteins govern their behavior. Curr Opin Struct Biol. 1996;6:11–17. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
