

Phosphorylation of AMPA receptors is required for sensory deprivation-induced homeostatic synaptic plasticity
- PMID: 21483826
- PMCID: PMC3069067
- DOI: 10.1371/journal.pone.0018264
Phosphorylation of AMPA receptors is required for sensory deprivation-induced homeostatic synaptic plasticity
Abstract
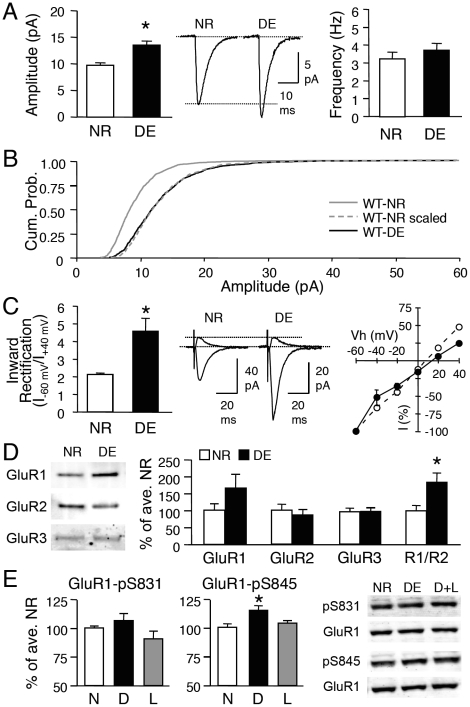
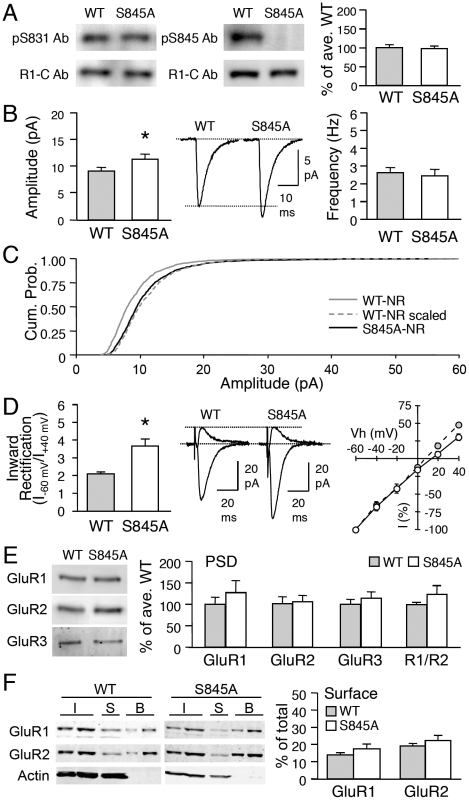
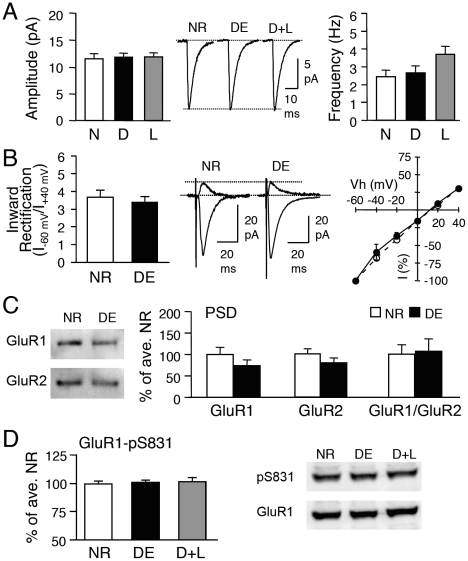
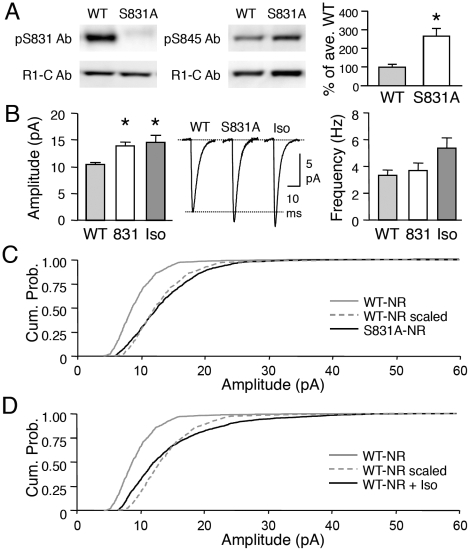
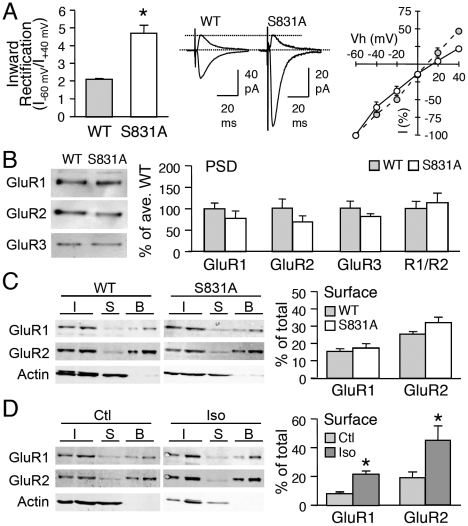
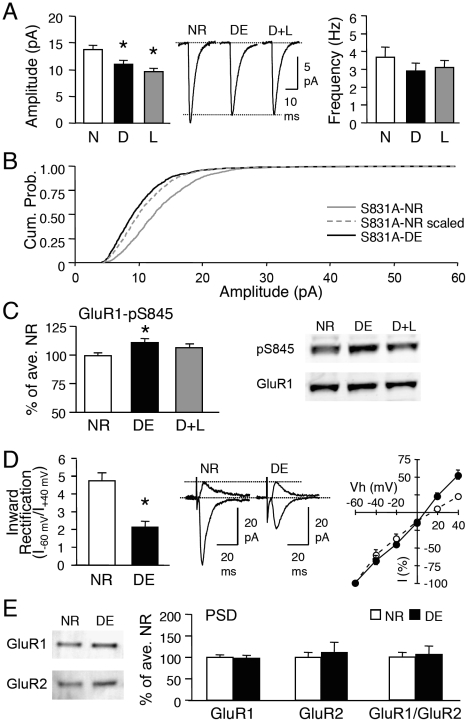
Sensory experience, and the lack thereof, can alter the function of excitatory synapses in the primary sensory cortices. Recent evidence suggests that changes in sensory experience can regulate the synaptic level of Ca(2+)-permeable AMPA receptors (CP-AMPARs). However, the molecular mechanisms underlying such a process have not been determined. We found that binocular visual deprivation, which is a well-established in vivo model to produce multiplicative synaptic scaling in visual cortex of juvenile rodents, is accompanied by an increase in the phosphorylation of AMPAR GluR1 (or GluA1) subunit at the serine 845 (S845) site and the appearance of CP-AMPARs at synapses. To address the role of GluR1-S845 in visual deprivation-induced homeostatic synaptic plasticity, we used mice lacking key phosphorylation sites on the GluR1 subunit. We found that mice specifically lacking the GluR1-S845 site (GluR1-S845A mutants), which is a substrate of cAMP-dependent kinase (PKA), show abnormal basal excitatory synaptic transmission and lack visual deprivation-induced homeostatic synaptic plasticity. We also found evidence that increasing GluR1-S845 phosphorylation alone is not sufficient to produce normal multiplicative synaptic scaling. Our study provides concrete evidence that a GluR1 dependent mechanism, especially S845 phosphorylation, is a necessary pre-requisite step for in vivo homeostatic synaptic plasticity.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
