

Herpes simplex virus dances with amyloid precursor protein while exiting the cell
- PMID: 21483850
- PMCID: PMC3069030
- DOI: 10.1371/journal.pone.0017966
Herpes simplex virus dances with amyloid precursor protein while exiting the cell
Abstract
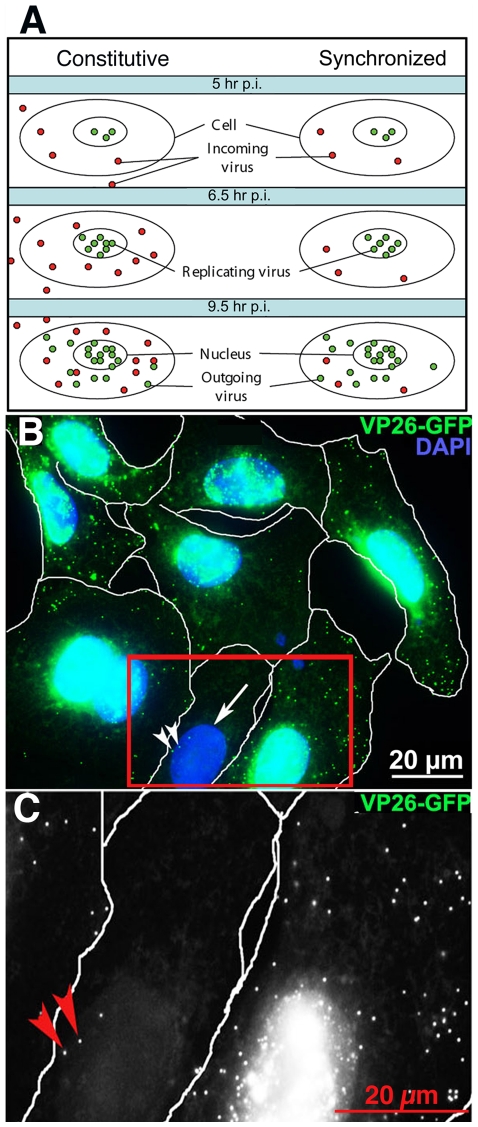
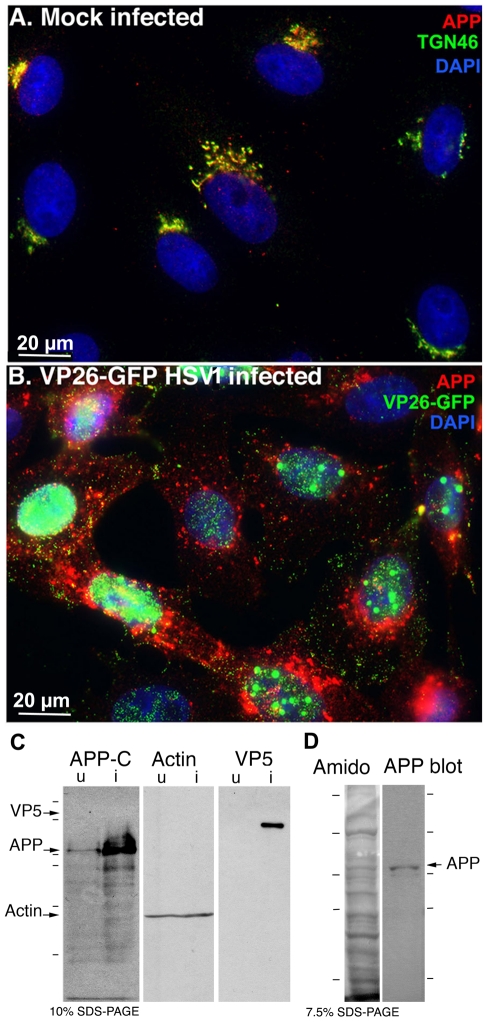
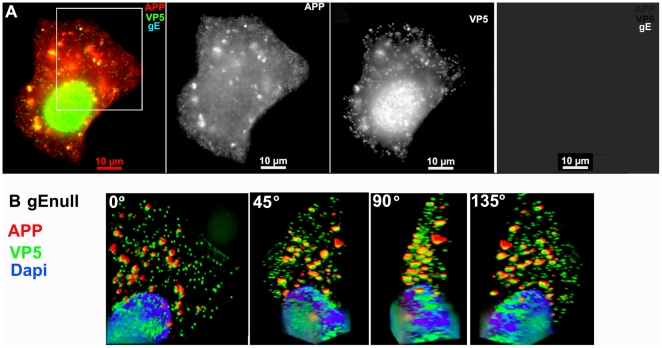
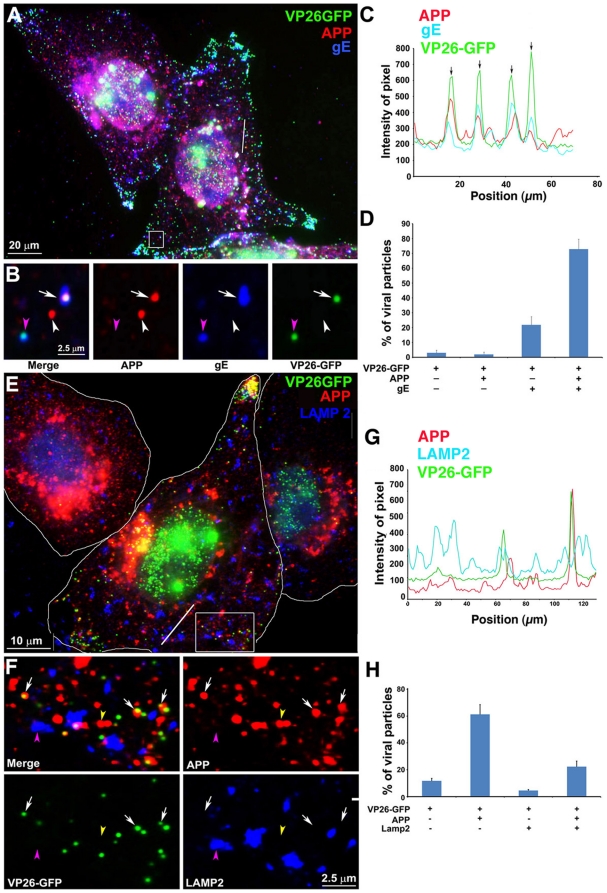
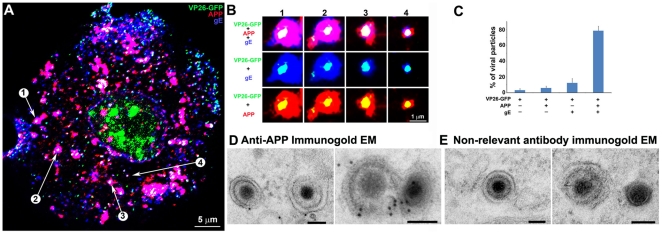
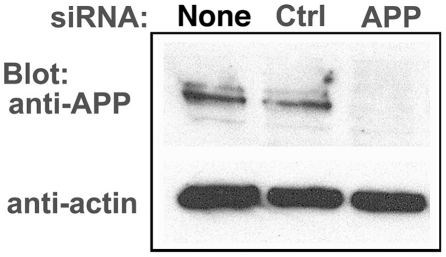
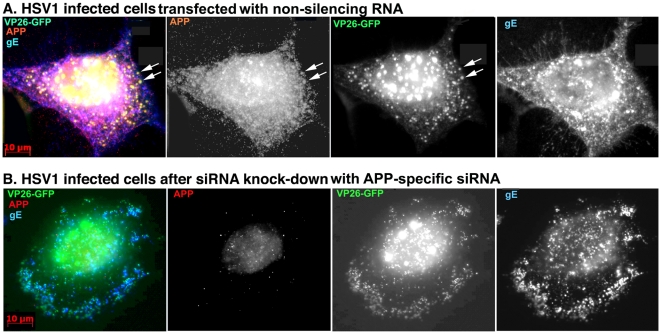
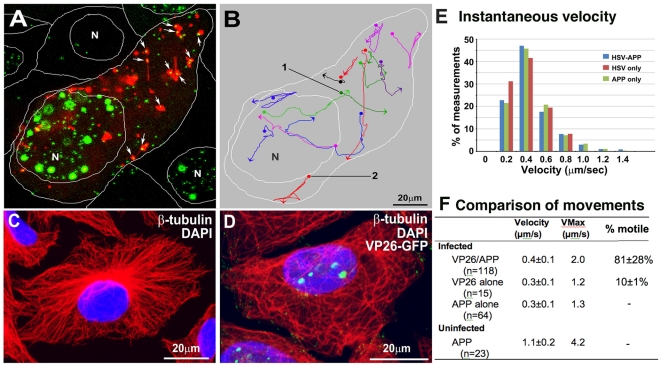
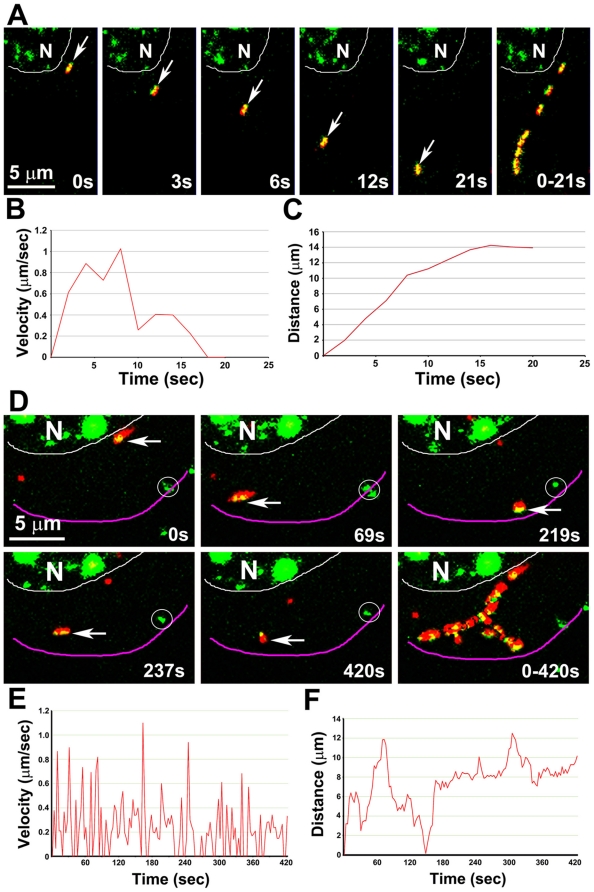
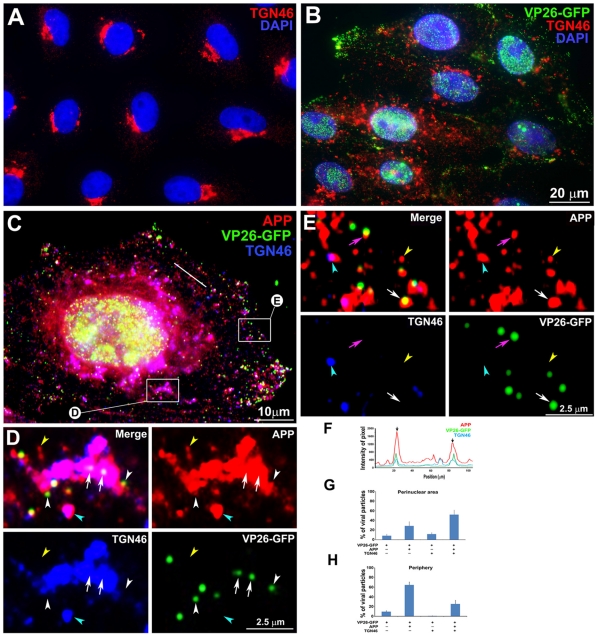
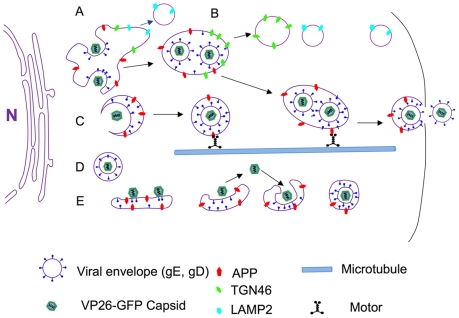
Herpes simplex type 1 (HSV1) replicates in epithelial cells and secondarily enters local sensory neuronal processes, traveling retrograde to the neuronal nucleus to enter latency. Upon reawakening newly synthesized viral particles travel anterograde back to the epithelial cells of the lip, causing the recurrent cold sore. HSV1 co-purifies with amyloid precursor protein (APP), a cellular transmembrane glycoprotein and receptor for anterograde transport machinery that when proteolyzed produces A-beta, the major component of senile plaques. Here we focus on transport inside epithelial cells of newly synthesized virus during its transit to the cell surface. We hypothesize that HSV1 recruits cellular APP during transport. We explore this with quantitative immuno-fluorescence, immuno-gold electron-microscopy and live cell confocal imaging. After synchronous infection most nascent VP26-GFP-labeled viral particles in the cytoplasm co-localize with APP (72.8+/-6.7%) and travel together with APP inside living cells (81.1+/-28.9%). This interaction has functional consequences: HSV1 infection decreases the average velocity of APP particles (from 1.1+/-0.2 to 0.3+/-0.1 µm/s) and results in APP mal-distribution in infected cells, while interplay with APP-particles increases the frequency (from 10% to 81% motile) and velocity (from 0.3+/-0.1 to 0.4+/-0.1 µm/s) of VP26-GFP transport. In cells infected with HSV1 lacking the viral Fc receptor, gE, an envelope glycoprotein also involved in viral axonal transport, APP-capsid interactions are preserved while the distribution and dynamics of dual-label particles differ from wild-type by both immuno-fluorescence and live imaging. Knock-down of APP with siRNA eliminates APP staining, confirming specificity. Our results indicate that most intracellular HSV1 particles undergo frequent dynamic interplay with APP in a manner that facilitates viral transport and interferes with normal APP transport and distribution. Such dynamic interactions between APP and HSV1 suggest a mechanistic basis for the observed clinical relationship between HSV1 seropositivity and risk of Alzheimer's disease.
Conflict of interest statement
Figures
References
-
- Lycke E, Kristensson K, Svennerholm B, Vahlne A, Ziegler R. Uptake and transport of herpes simplex virus in neurites of rat dorsal root ganglia cells in culture. J Gen Virol. 1984;65(Pt 1):55–64. - PubMed
-
- Kristensson K, Lycke E, Roytta M, Svennerholm B, Vahlne A. Neuritic transport of herpes simplex virus in rat sensory neurons in vitro. Effects of substances interacting with microtubular function and axonal flow [nocodazole, taxol and erythro-9-3-(2-hydroxynonyl)adenine]. J Gen Virol. 1986;67(Pt 9):2023–2028. - PubMed
-
- Enquist LW, Tomishima MJ, Gross S, Smith GA. Directional spread of an alpha-herpesvirus in the nervous system. Vet Microbiol. 2002;86:5–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
