

Loss of olfactory cell adhesion molecule reduces the synchrony of mitral cell activity in olfactory glomeruli
- PMID: 21486802
- PMCID: PMC3090595
- DOI: 10.1113/jphysiol.2011.206276
Loss of olfactory cell adhesion molecule reduces the synchrony of mitral cell activity in olfactory glomeruli
Abstract
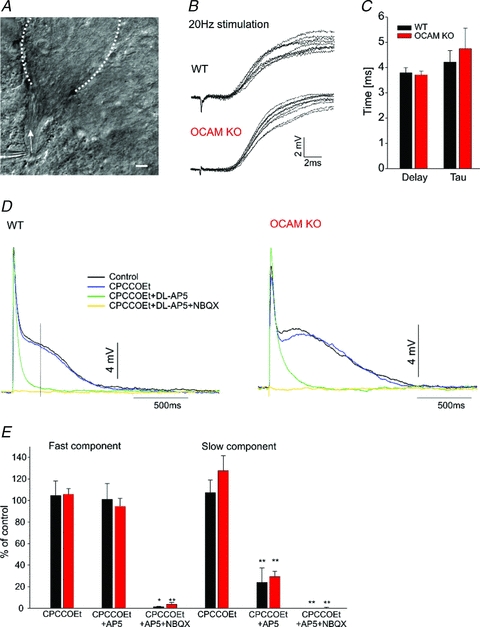
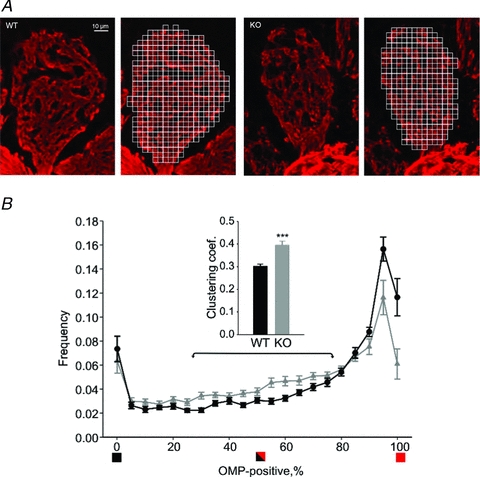
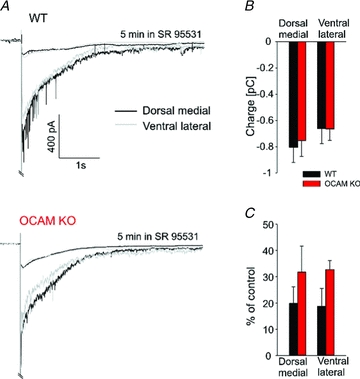
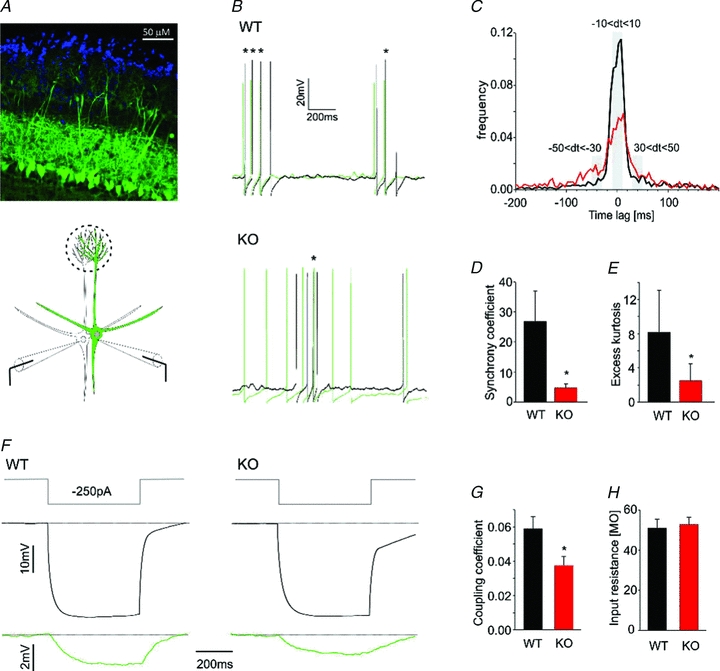
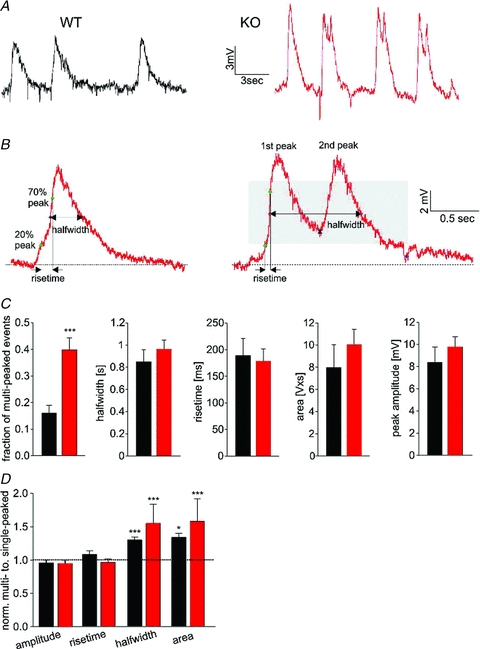
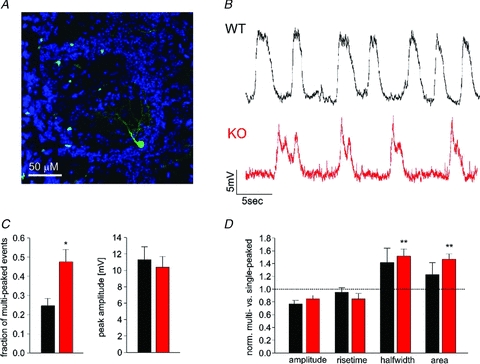
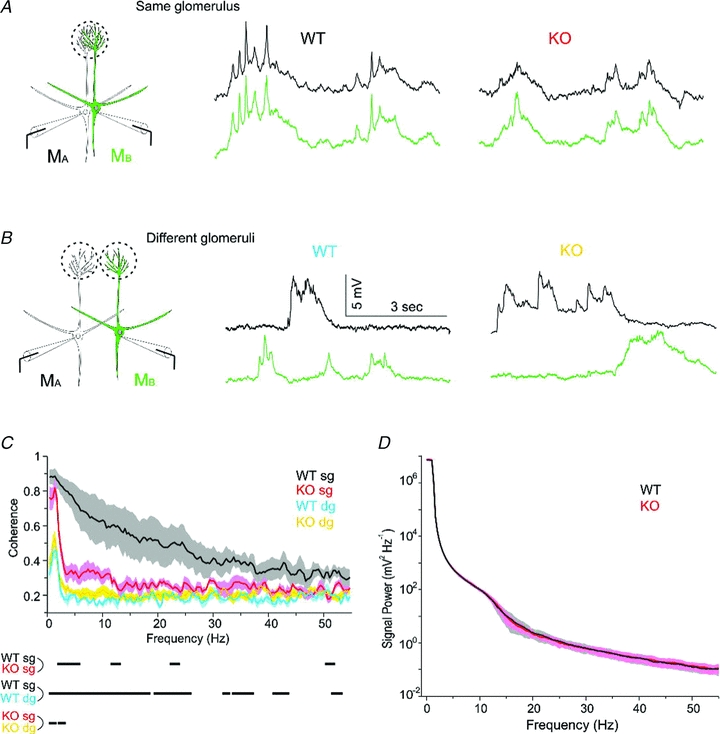
Odours generate activity in olfactory receptor neurons, whose axons contact the dendritic tufts of mitral cells within olfactory bulb glomeruli. These axodendritic synapses are anatomically separated from dendrodendritic synapses within each glomerulus. Mitral cells within a glomerulus show highly synchronized activity as assessed with whole-cell recording from pairs of mitral cells. We examined glomerular activity in mice lacking the olfactory cell adhesion molecule (OCAM). Glomeruli in mice lacking OCAM show a redistribution of synaptic subcompartments, but the total area occupied by axonal inputs was similar to wild-type mice. Stimulation of olfactory nerve bundles showed that excitatory synaptic input to mitral cells as well as dendrodendritic inhibition was unaffected in the knockout. However, correlated spiking in mitral cells was significantly reduced, as was electrical coupling between apical dendrites. To analyse slow network dynamics we induced slow oscillations with a glutamate uptake blocker. Evoked and spontaneous slow oscillations in mitral cells and external tufted cells were broader and had multiple peaks in OCAM knockout mice, indicating that synchrony of slow glomerular activity was also reduced. To assess the degree of shared activity between mitral cells under physiological conditions, we analysed spontaneous sub-threshold voltage oscillations using coherence analysis. Coherent activity was markedly reduced in cells from OCAM knockout mice across a broad range of frequencies consistent with a decrease in tightly time-locked activity. We suggest that synchronous activity within each glomerulus is dependent on segregation of synaptic subcompartments.
Figures
References
-
- Abraham NM, Spors H, Carleton A, Margrie TW, Kuner T, Schaefer AT. Maintaining accuracy at the expense of speed: stimulus similarity defines odor discrimination time in mice. Neuron. 2004;44:865–876. - PubMed
-
- Baker H, Grillo M, Margolis FL. Biochemical and immunocytochemical characterization of olfactory marker protein in the rodent central nervous system. J Comp Neurol. 1989;285:246–261. - PubMed
-
- Briggman KL, Denk W. Towards neural circuit reconstruction with volume electron microscopy techniques. Curr Opin Neurobiol. 2006;16:562–570. - PubMed
-
- Buck L, Axel R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 1991;65:175–187. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases