

Local mobile gene pools rapidly cross species boundaries to create endemicity within global Vibrio cholerae populations
- PMID: 21486909
- PMCID: PMC3073641
- DOI: 10.1128/mBio.00335-10
Local mobile gene pools rapidly cross species boundaries to create endemicity within global Vibrio cholerae populations
Abstract
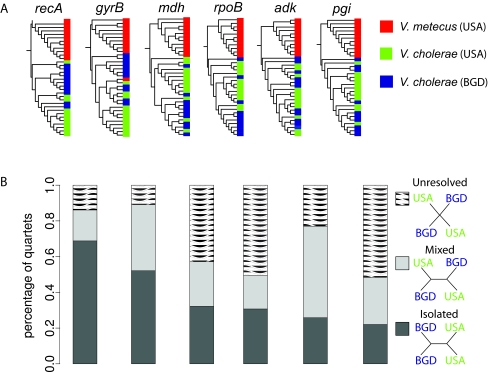
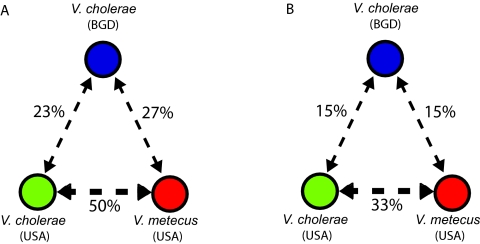
Vibrio cholerae represents both an environmental pathogen and a widely distributed microbial species comprised of closely related strains occurring in the tropical to temperate coastal ocean across the globe (Colwell RR, Science 274:2025-2031, 1996; Griffith DC, Kelly-Hope LA, Miller MA, Am. J. Trop. Med. Hyg. 75:973-977, 2006; Reidl J, Klose KE, FEMS Microbiol. Rev. 26:125-139, 2002). However, although this implies dispersal and growth across diverse environmental conditions, how locally successful populations assemble from a possibly global gene pool, relatively unhindered by geographic boundaries, remains poorly understood. Here, we show that environmental Vibrio cholerae possesses two, largely distinct gene pools: one is vertically inherited and globally well mixed, and the other is local and rapidly transferred across species boundaries to generate an endemic population structure. While phylogeographic analysis of isolates collected from Bangladesh and the U.S. east coast suggested strong panmixis for protein-coding genes, there was geographic structure in integrons, which are the only genomic islands present in all strains of V. cholerae (Chun J, et al., Proc. Natl. Acad. Sci. U. S. A. 106:15442-15447, 2009) and are capable of acquiring and expressing mobile gene cassettes. Geographic differentiation in integrons arises from high gene turnover, with acquisition from a locally co-occurring sister species being up to twice as likely as exchange with conspecific but geographically distant V. cholerae populations. IMPORTANCE Functional predictions of integron genes show the predominance of secondary metabolism and cell surface modification, which is consistent with a role in competition and predation defense. We suggest that the integron gene pool's distinctness and tempo of sharing are adaptive in allowing rapid conversion of genomes to reflect local ecological constraints. Because the integron is frequently the main element differentiating clinical strains (Chun J, et al., Proc. Natl. Acad. Sci. U. S. A. 106:15442-15447, 2009) and its recombinogenic activity is directly stimulated by environmental stresses (Guerin E, et al., Science 324:1034, 2009), these observations are relevant for local emergence and subsequent dispersal.
Figures


References
-
- Colwell RR. 1996. Global climate and infectious disease: the cholera paradigm. Science 274:2025–2031 - PubMed
-
- Lapierre P, Gogarten JP. 2009. Estimating the size of the bacterial pan-genome. Trends Genet. 25:107–110 - PubMed
-
- Tettelin H, Riley D, Cattuto C, Medini D. 2008. Comparative genomics: the bacterial pan-genome. Curr. Opin. Microbiol. 12:472–477 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
