

Truncation of monocyte chemoattractant protein 1 by plasmin promotes blood-brain barrier disruption
- PMID: 21486949
- PMCID: PMC3078815
- DOI: 10.1242/jcs.082834
Truncation of monocyte chemoattractant protein 1 by plasmin promotes blood-brain barrier disruption
Abstract
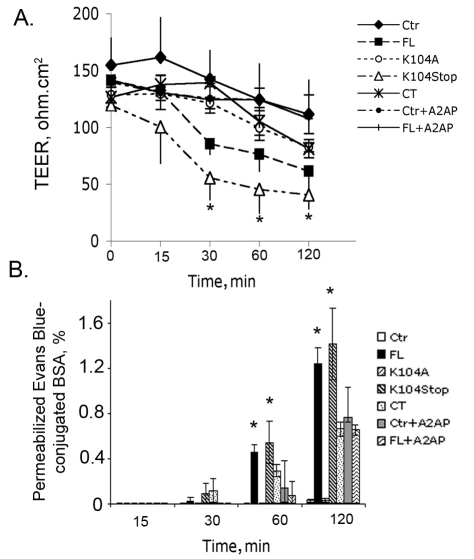
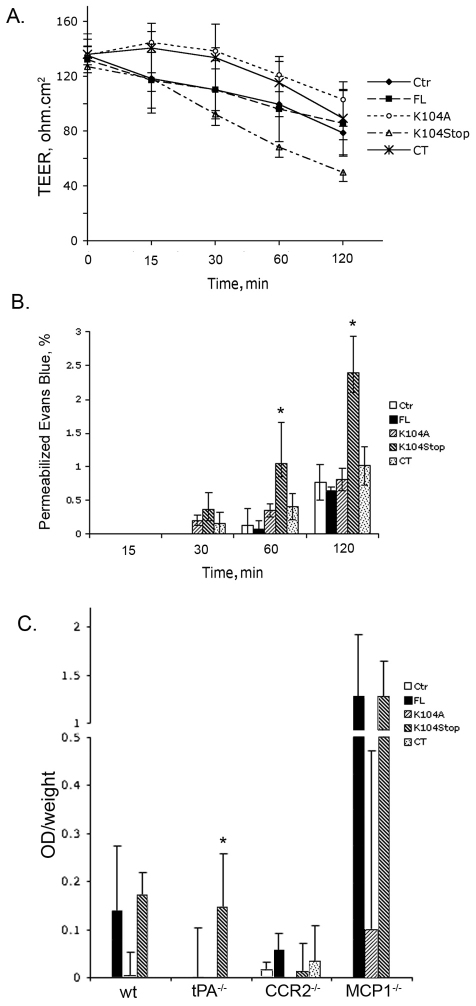
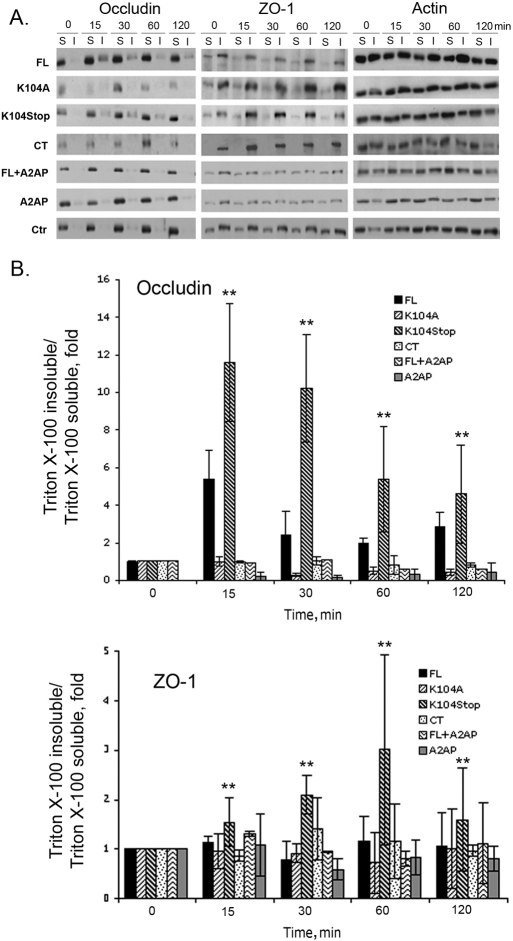
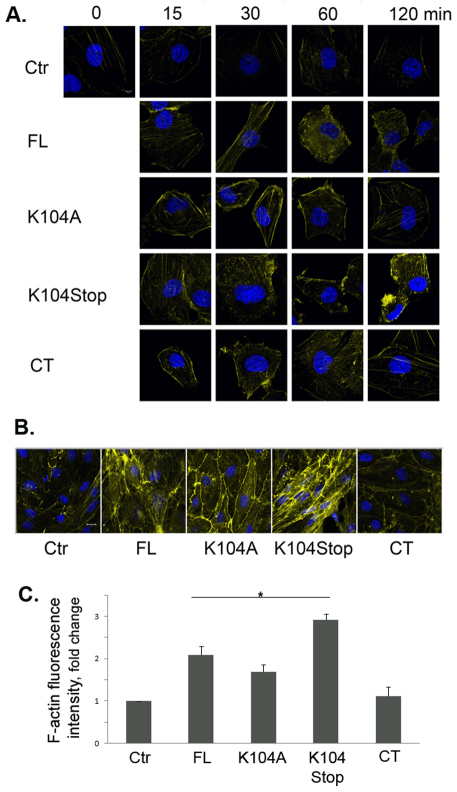
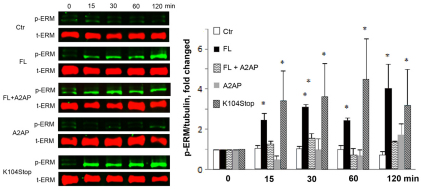
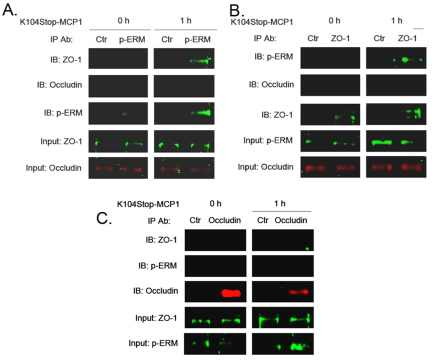
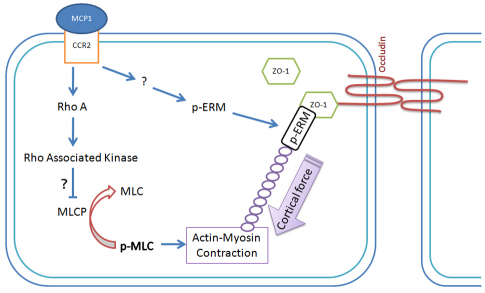
Previous studies have shown that plasmin cleaves monocyte chemoattractant protein 1 (MCP1; officially known as C-C motif chemokine 2, CCL2) at K104, and this cleavage enhances its chemotactic potency significantly. Accumulating evidence reveals that MCP1 also disrupts the integrity of the blood-brain barrier (BBB). Here, we show that K104Stop-MCP1, truncated at the K104 where plasmin would normally cleave, is more efficient than the full-length protein (FL-MCP1) in compromising the integrity of the BBB in in vitro and in vivo models. K104Stop-MCP1 increases the permeability of BBB in both wild-type mice and mice deficient for tissue plasminogen activator (tPA), which converts plasminogen into active plasmin, suggesting that plasmin-mediated truncation of MCP1 plays an important role in BBB compromise. Furthermore, we show that the mechanisms underlying MCP1-induced BBB disruption involve redistribution of tight junction proteins (occludin and ZO-1) and reorganization of the actin cytoskeleton. Finally, we show that the redistribution of ZO-1 is mediated by phosphorylation of ezrin-radixin-moesin (ERM) proteins. These findings identify plasmin as a key signaling molecule in the regulation of BBB integrity and suggest that plasmin inhibitors might be used to modulate diseases accompanied by BBB compromise.
Figures

References
-
- Ahdieh M., Vandenbos T., Youakim A. (2001). Lung epithelial barrier function and wound healing are decreased by IL-4 and IL-13 and enhanced by IFN-gamma. Am. J. Physiol. Cell Physiol. 281, C2029-C2038 - PubMed
-
- Alter A., Duddy M., Hebert S., Biernacki K., Prat A., Antel J. P., Yong V. W., Nuttall R. K., Pennington C. J., Edwards D. R., et al. (2003). Determinants of human B cell migration across brain endothelial cells. J. Immunol. 170, 4497-4505 - PubMed
-
- Bretscher A., Gary R., Berryman M. (1995). Soluble ezrin purified from placenta exists as stable monomers and elongated dimers with masked C-terminal ezrin-radixin-moesin association domains. Biochemistry 34, 16830-16837 - PubMed
-
- Calabria A. R., Weidenfeller C., Jones A. R., de Vries H. E., Shusta E. V. (2006). Puromycin-purified rat brain microvascular endothelial cell cultures exhibit improved barrier properties in response to glucocorticoid induction. J. Neurochem. 97, 922-933 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
