

The Bphi008a gene interacts with the ethylene pathway and transcriptionally regulates MAPK genes in the response of rice to brown planthopper feeding
- PMID: 21487048
- PMCID: PMC3177281
- DOI: 10.1104/pp.111.174334
The Bphi008a gene interacts with the ethylene pathway and transcriptionally regulates MAPK genes in the response of rice to brown planthopper feeding
Abstract
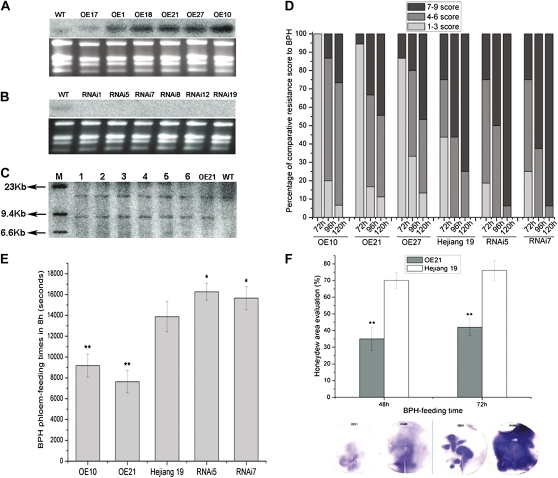
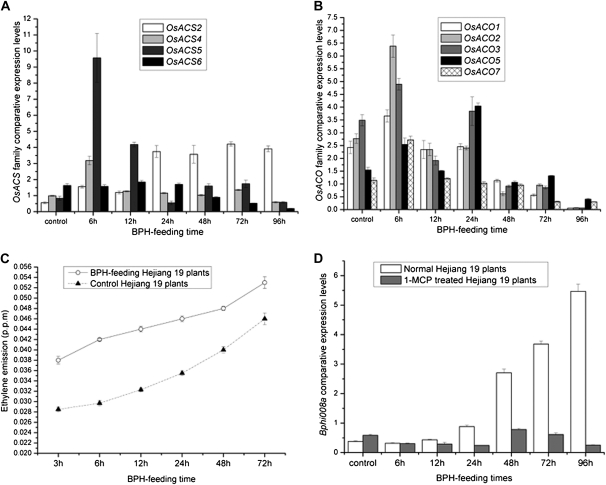
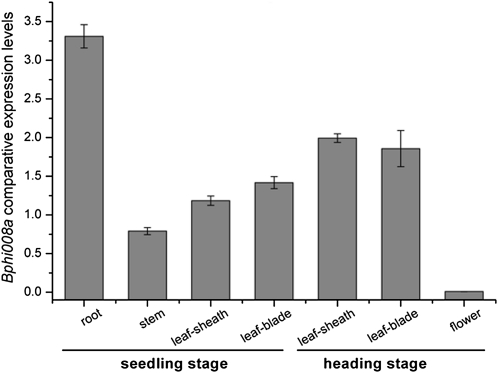
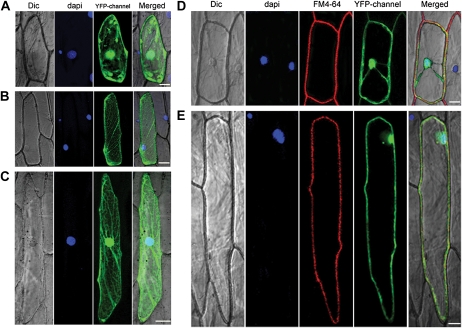
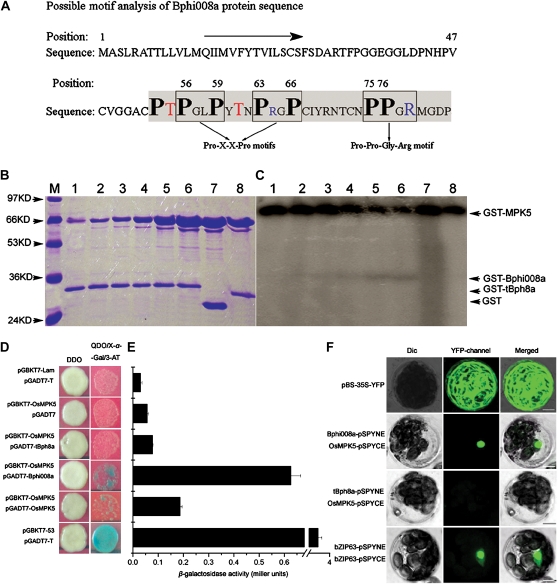
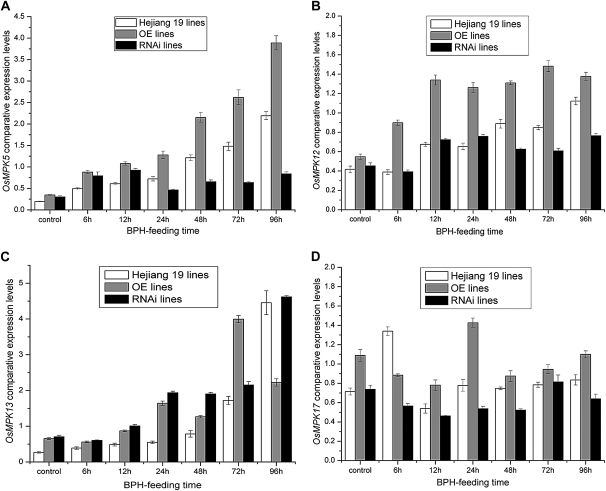
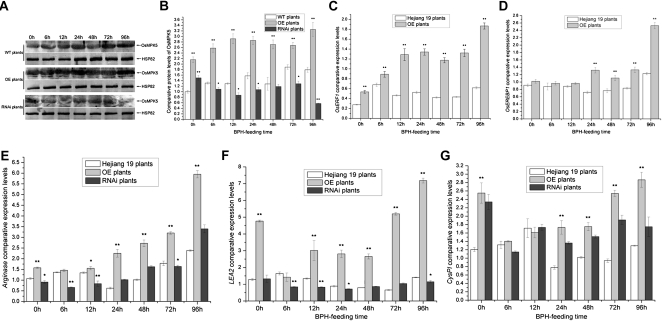
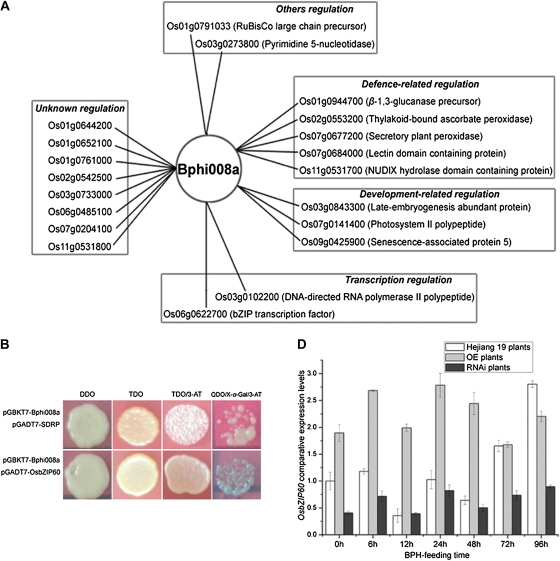
We examined ways in which the Brown planthopper induced008a (Bphi008a; AY256682) gene of rice (Oryza sativa) enhances the plant's resistance to a specialist herbivore, the brown planthopper (BPH; Nilaparvata lugens). Measurement of the expression levels of ethylene synthases and of ethylene emissions showed that BPH feeding rapidly initiated the ethylene signaling pathway and up-regulated Bphi008a transcript levels after 6 to 96 h of feeding. In contrast, blocking ethylene transduction (using 1-methylcyclopropene) reduced Bphi008a transcript levels in wild-type plants fed upon by BPH. In vitro kinase assays showed that Bphi008a can be phosphorylated by rice Mitogen-activated Protein Kinase5 (OsMPK5), and yeast two-hybrid assays demonstrated that the carboxyl-terminal proline-rich region of Bphi008a interacts directly with this kinase. Furthermore, bimolecular fluorescence complementation assays showed that this interaction occurs in the nucleus. Subsequently, we found that Bphi008a up-regulation and down-regulation were accompanied by different changes in transcription levels of OsMPK5, OsMPK12, OsMPK13, and OsMPK17 in transgenic plants. Immunoblot analysis also showed that the OsMPK5 protein level increased in overexpressing plants and decreased in RNA interference plants after BPH feeding. In transgenic lines, changes in the expression levels of several enzymes that are important components of the defenses against the BPH were also observed. Finally, yeast two-hybrid screening results showed that Bphi008a is able to interact with a b-ZIP transcription factor (OsbZIP60) and a RNA polymerase polypeptide (SDRP).
Figures

References
-
- Andersen CL, Jensen JL, Ørntoft TF. (2004) Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res 64: 5245–5250 - PubMed
-
- Asai T, Tena G, Plotnikova J, Willmann MR, Chiu WL, Gomez-Gomez L, Boller T, Ausubel FM, Sheen J. (2002) MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415: 977–983 - PubMed
-
- Ausubel FM. (2005) Are innate immune signaling pathways in plants and animals conserved? Nat Immunol 6: 973–979 - PubMed
-
- Baldwin IT, Kessler A, Halitschke R. (2002) Volatile signaling in plant-plant-herbivore interactions: what is real? Curr Opin Plant Biol 5: 351–354 - PubMed
-
- Berrocal-Lobo M, Molina A, Solano R. (2002) Constitutive expression of ETHYLENE-RESPONSE-FACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi. Plant J 29: 23–32 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
