

Discovery of permuted and recently split transfer RNAs in Archaea
- PMID: 21489296
- PMCID: PMC3218864
- DOI: 10.1186/gb-2011-12-4-r38
Discovery of permuted and recently split transfer RNAs in Archaea
Abstract
Background: As in eukaryotes, precursor transfer RNAs in Archaea often contain introns that are removed in tRNA maturation. Two unrelated archaeal species display unique pre-tRNA processing complexity in the form of split tRNA genes, in which two to three segments of tRNAs are transcribed from different loci, then trans-spliced to form a mature tRNA. Another rare type of pre-tRNA, found only in eukaryotic algae, is permuted, where the 3' half is encoded upstream of the 5' half, and must be processed to be functional.
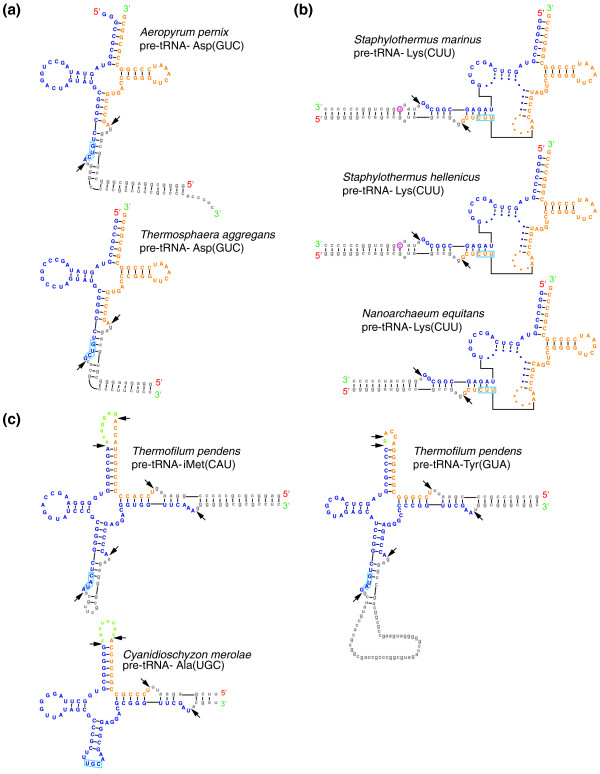
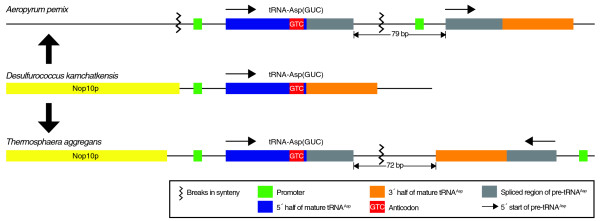
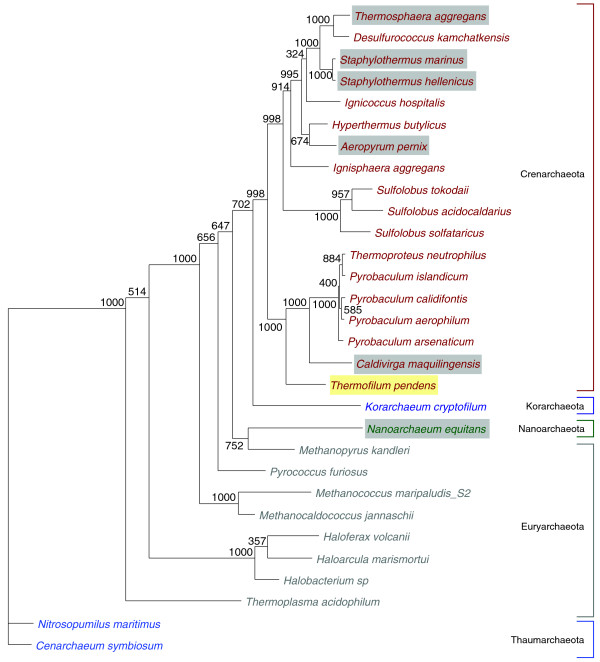
Results: Using an improved version of the gene-finding program tRNAscan-SE, comparative analyses and experimental verifications, we have now identified four novel trans-spliced tRNA genes, each in a different species of the Desulfurococcales branch of the Archaea: tRNA(Asp(GUC)) in Aeropyrum pernix and Thermosphaera aggregans, and tRNA(Lys(CUU)) in Staphylothermus hellenicus and Staphylothermus marinus. Each of these includes features surprisingly similar to previously studied split tRNAs, yet comparative genomic context analysis and phylogenetic distribution suggest several independent, relatively recent splitting events. Additionally, we identified the first examples of permuted tRNA genes in Archaea: tRNA(iMet(CAU)) and tRNA(Tyr(GUA)) in Thermofilum pendens, which appear to be permuted in the same arrangement seen previously in red alga.
Conclusions: Our findings illustrate that split tRNAs are sporadically spread across a major branch of the Archaea, and that permuted tRNAs are a new shared characteristic between archaeal and eukaryotic species. The split tRNA discoveries also provide new clues to their evolutionary history, supporting hypotheses for recent acquisition via viral or other mobile elements.
© 2011 Chan et al.; licensee BioMed Central Ltd.
Figures

Similar articles
-
Disrupted tRNA gene diversity and possible evolutionary scenarios.J Mol Evol. 2009 Nov;69(5):497-504. doi: 10.1007/s00239-009-9294-6. Epub 2009 Oct 14. J Mol Evol. 2009. PMID: 19826747
-
Exploring tRNA gene cluster in archaea.Mem Inst Oswaldo Cruz. 2019 Jan 7;114:e180348. doi: 10.1590/0074-02760180348. Mem Inst Oswaldo Cruz. 2019. PMID: 30624459 Free PMC article.
-
SPLITS: a new program for predicting split and intron-containing tRNA genes at the genome level.In Silico Biol. 2006;6(5):411-8. In Silico Biol. 2006. PMID: 17274770
-
Archaeal aminoacyl-tRNA synthesis: diversity replaces dogma.Genetics. 1999 Aug;152(4):1269-76. doi: 10.1093/genetics/152.4.1269. Genetics. 1999. PMID: 10430557 Free PMC article. Review.
-
Archaeal introns: splicing, intercellular mobility and evolution.Trends Biochem Sci. 1997 Sep;22(9):326-31. doi: 10.1016/s0968-0004(97)01113-4. Trends Biochem Sci. 1997. PMID: 9301331 Review.
Cited by
-
Experimental confirmation of a whole set of tRNA molecules in two archaeal species.Int J Mol Sci. 2015 Jan 20;16(1):2187-203. doi: 10.3390/ijms16012187. Int J Mol Sci. 2015. PMID: 25608653 Free PMC article.
-
Dimerization confers increased stability to nucleases in 5' halves from glycine and glutamic acid tRNAs.Nucleic Acids Res. 2018 Sep 28;46(17):9081-9093. doi: 10.1093/nar/gky495. Nucleic Acids Res. 2018. PMID: 29893896 Free PMC article.
-
Circularly permuted tRNA genes: their expression and implications for their physiological relevance and development.Front Genet. 2014 Apr 1;5:63. doi: 10.3389/fgene.2014.00063. eCollection 2014. Front Genet. 2014. PMID: 24744771 Free PMC article. Review.
-
A Comprehensive tRNA Genomic Survey Unravels the Evolutionary History of tRNA Arrays in Prokaryotes.Genome Biol Evol. 2015 Dec 28;8(1):282-95. doi: 10.1093/gbe/evv254. Genome Biol Evol. 2015. PMID: 26710853 Free PMC article.
-
Structural Basis for tRNA Mimicry by a Bacterial Y RNA.Structure. 2018 Dec 4;26(12):1635-1644.e3. doi: 10.1016/j.str.2018.09.001. Epub 2018 Oct 11. Structure. 2018. PMID: 30318468 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
