

Toward mechanistic classification of enzyme functions
- PMID: 21489855
- PMCID: PMC3551611
- DOI: 10.1016/j.cbpa.2011.03.008
Toward mechanistic classification of enzyme functions
Abstract
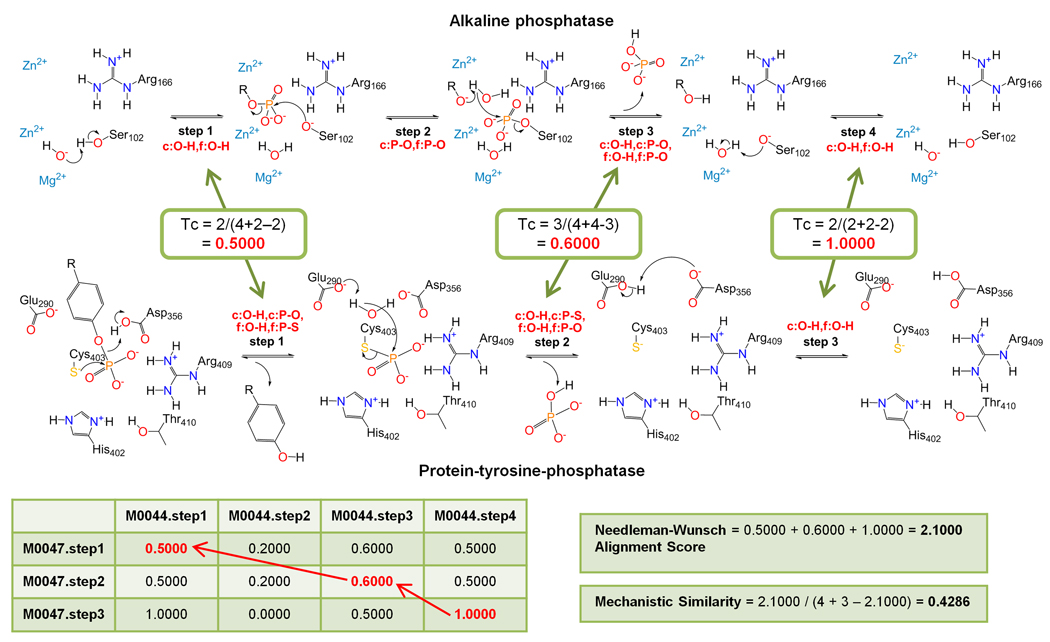
Classification of enzyme function should be quantitative, computationally accessible, and informed by sequences and structures to enable use of genomic information for functional inference and other applications. Large-scale studies have established that divergently evolved enzymes share conserved elements of structure and common mechanistic steps and that convergently evolved enzymes often converge to similar mechanisms too, suggesting that reaction mechanisms could be used to develop finer-grained functional descriptions than provided by the Enzyme Commission (EC) system currently in use. Here we describe how evolution informs these structure-function mappings and review the databases that store mechanisms of enzyme reactions along with recent developments to measure ligand and mechanistic similarities. Together, these provide a foundation for new classifications of enzyme function.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- IUBMB. Enzyme Nomenclature 1992: Recommendations of the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology on the nomenclature and classification of enzymes. Academic Press; 1992.
-
- Babbitt PC. Definitions of enzyme function for the structural genomics era. Curr Opin Chem Biol. 2003;7:230–237. - PubMed
-
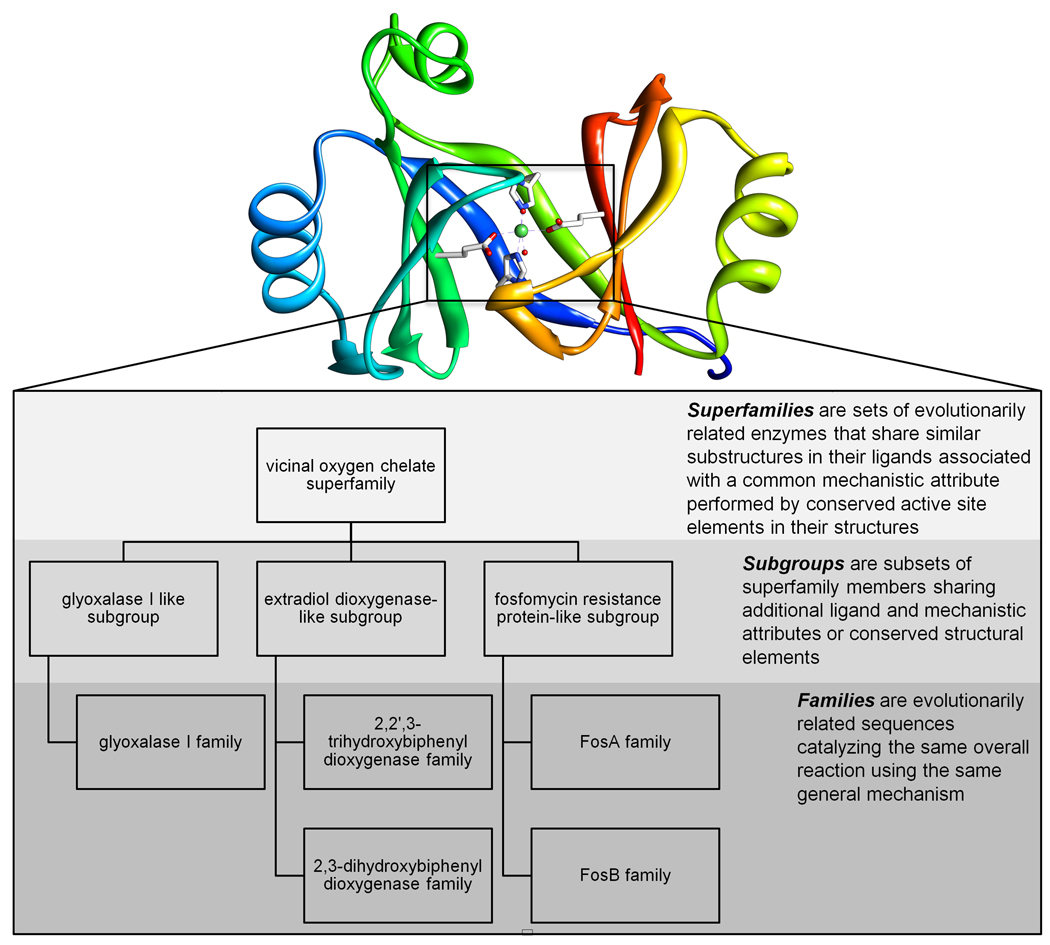
- Gerlt JA, Babbitt PC. Divergent evolution of enzymatic function: mechanistically diverse superfamilies and functionally distinct suprafamilies. Annu Rev Biochem. 2001;70:209–246. - PubMed
-
- Glasner ME, Gerlt JA, Babbitt PC. Evolution of enzyme superfamilies. Curr Opin Chem Biol. 2006;10:492–497. - PubMed
-
-
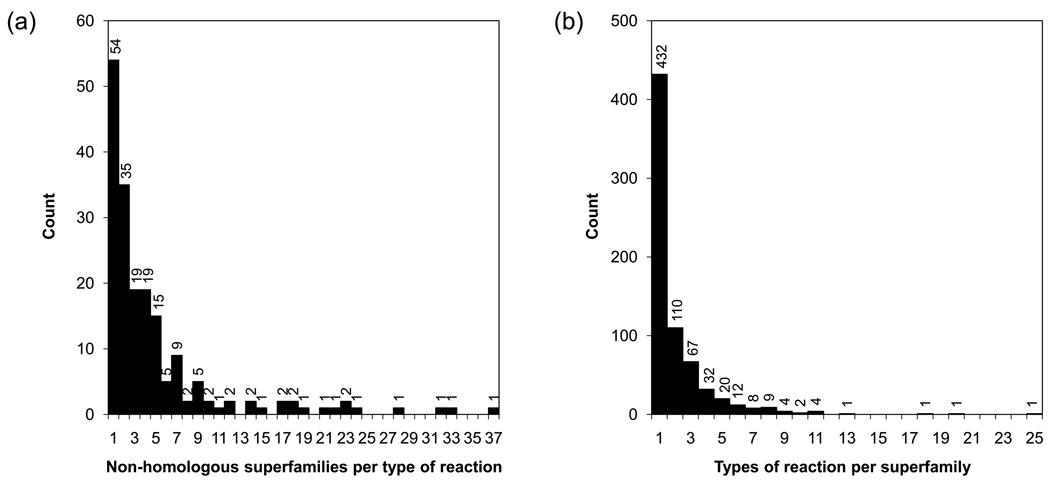
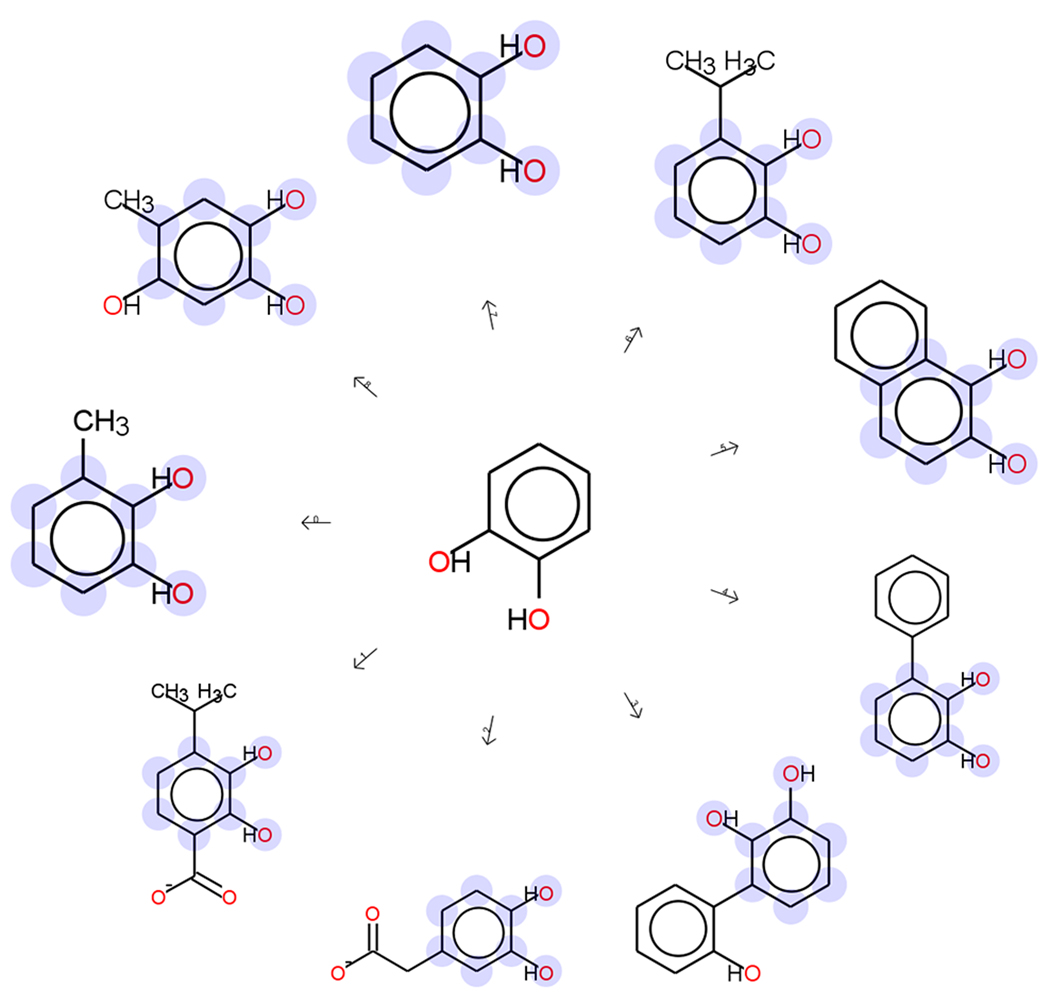
Omelchenko MV, Galperin MY, Wolf YI, Koonin EV. Non-homologous isofunctional enzymes: a systematic analysis of alternative solutions in enzyme evolution. Biol Direct. 2010;5:31. This is an updated survey of convergently evolved enzymes that identifies 186 groups of non-homologous enzymes that catalyze identical chemical transformations.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
