

Direct membrane association drives mitochondrial fission by the Parkinson disease-associated protein alpha-synuclein
- PMID: 21489994
- PMCID: PMC3121472
- DOI: 10.1074/jbc.M110.213538
Direct membrane association drives mitochondrial fission by the Parkinson disease-associated protein alpha-synuclein
Abstract
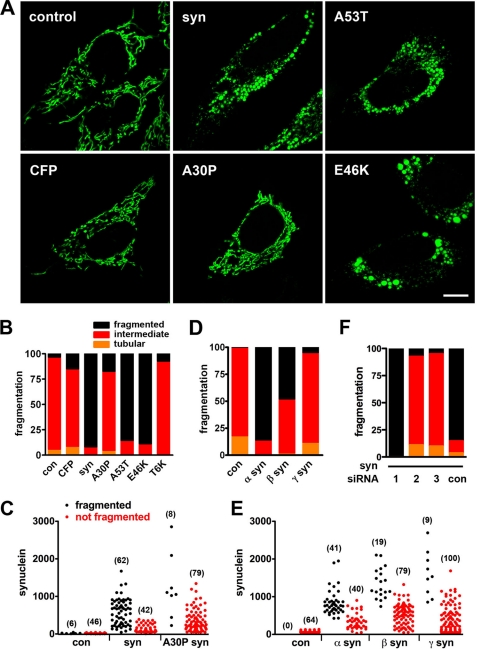
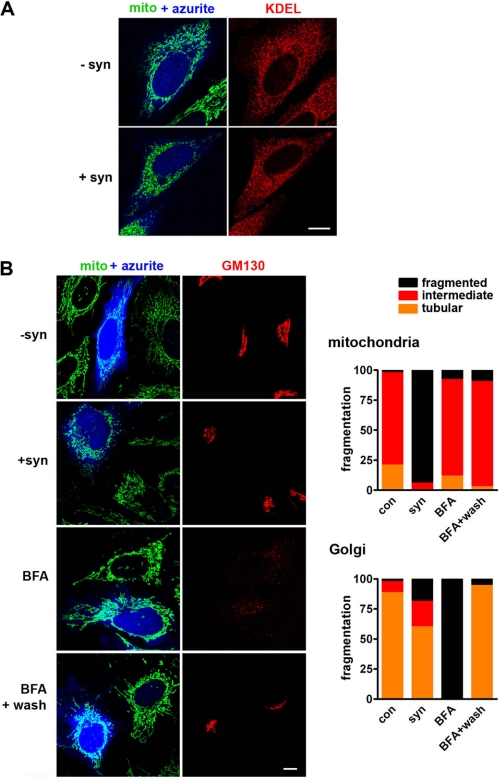
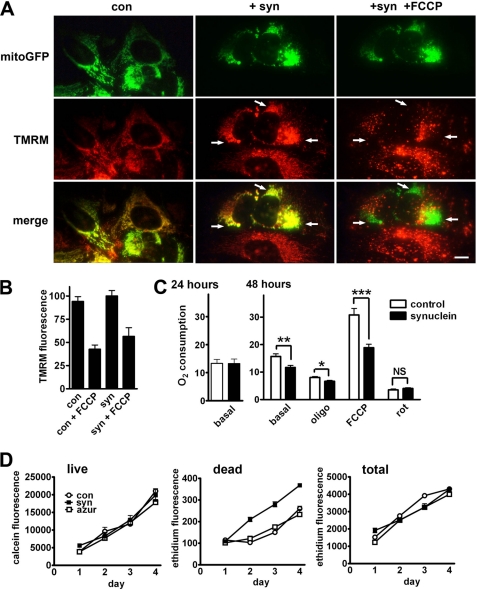
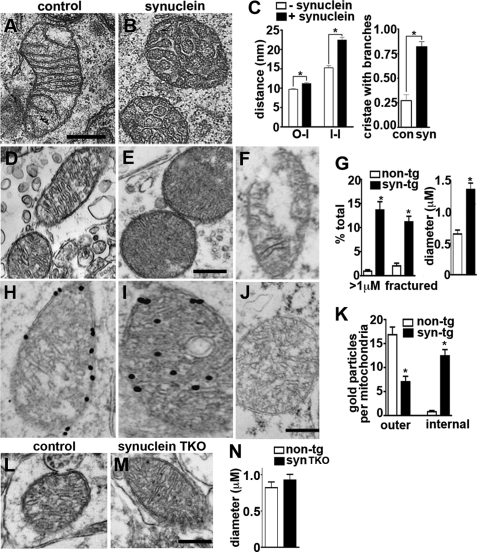
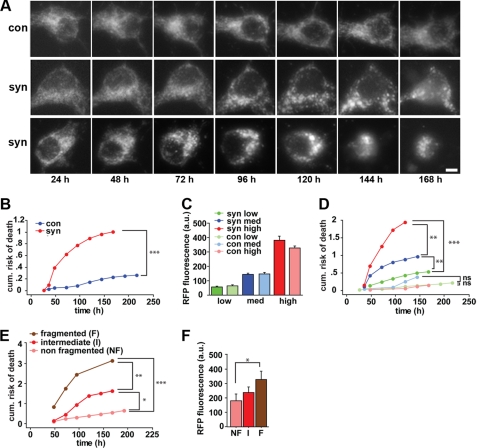
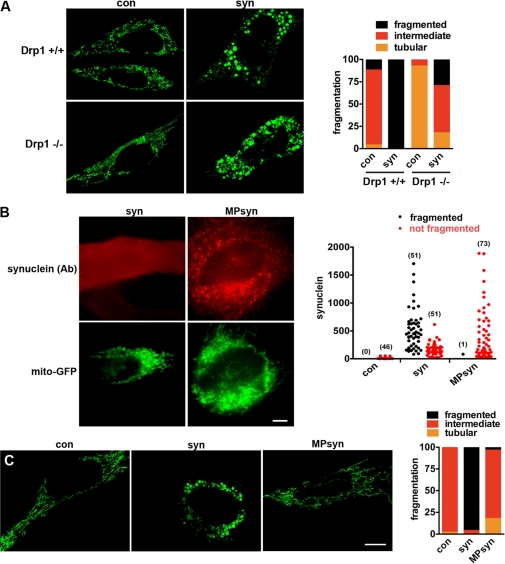
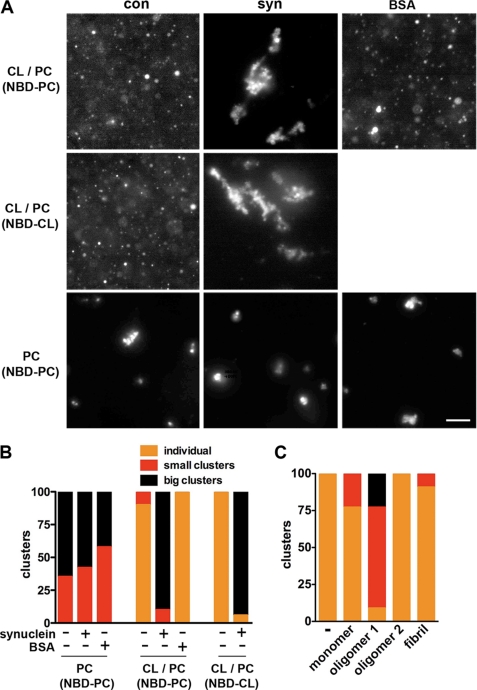
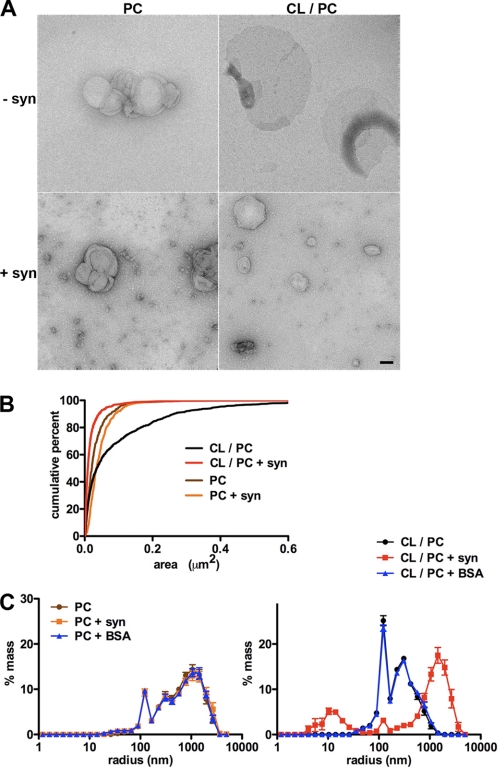
The protein α-synuclein has a central role in Parkinson disease, but the mechanism by which it contributes to neural degeneration remains unknown. We now show that the expression of α-synuclein in mammalian cells, including neurons in vitro and in vivo, causes the fragmentation of mitochondria. The effect is specific for synuclein, with more fragmentation by α- than β- or γ-isoforms, and it is not accompanied by changes in the morphology of other organelles or in mitochondrial membrane potential. However, mitochondrial fragmentation is eventually followed by a decline in respiration and neuronal death. The fragmentation does not require the mitochondrial fission protein Drp1 and involves a direct interaction of synuclein with mitochondrial membranes. In vitro, synuclein fragments artificial membranes containing the mitochondrial lipid cardiolipin, and this effect is specific for the small oligomeric forms of synuclein. α-Synuclein thus exerts a primary and direct effect on the morphology of an organelle long implicated in the pathogenesis of Parkinson disease.
Figures
References
-
- Schapira A. H., Mann V. M., Cooper J. M., Dexter D., Daniel S. E., Jenner P., Clark J. B., Marsden C. D. (1990) J. Neurochem. 55, 2142–2145 - PubMed
-
- Bender A., Krishnan K. J., Morris C. M., Taylor G. A., Reeve A. K., Perry R. H., Jaros E., Hersheson J. S., Betts J., Klopstock T., Taylor R. W., Turnbull D. M. (2006) Nat. Genet. 38, 515–517 - PubMed
-
- Valente E. M., Abou-Sleiman P. M., Caputo V., Muqit M. M., Harvey K., Gispert S., Ali Z., Del Turco D., Bentivoglio A. R., Healy D. G., Albanese A., Nussbaum R., González-Maldonado R., Deller T., Salvi S., Cortelli P., Gilks W. P., Latchman D. S., Harvey R. J., Dallapiccola B., Auburger G., Wood N. W. (2004) Science 304, 1158–1160 - PubMed
-
- Polymeropoulos M. H., Lavedan C., Leroy E., Ide S. E., Dehejia A., Dutra A., Pike B., Root H., Rubenstein J., Boyer R., Stenroos E. S., Chandrasekharappa S., Athanassiadou A., Papapetropoulos T., Johnson W. G., Lazzarini A. M., Duvoisin R. C., Di Iorio G., Golbe L. I., Nussbaum R. L. (1997) Science 276, 2045–2047 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01AG022074/AG/NIA NIH HHS/United States
- NS057096/NS/NINDS NIH HHS/United States
- R01 NS039074/NS/NINDS NIH HHS/United States
- GM089853/GM/NIGMS NIH HHS/United States
- P30 NS057096/NS/NINDS NIH HHS/United States
- R01 AG018440/AG/NIA NIH HHS/United States
- 2R01NS39074/NS/NINDS NIH HHS/United States
- AG022074/AG/NIA NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R01 GM089853/GM/NIGMS NIH HHS/United States
- K08 NS062954/NS/NINDS NIH HHS/United States
- P01 DA010154/DA/NIDA NIH HHS/United States
- DA10154/DA/NIDA NIH HHS/United States
- 1KO8NS062954-01A1/NS/NINDS NIH HHS/United States
- R01 NS062715/NS/NINDS NIH HHS/United States
- R37 AG018440/AG/NIA NIH HHS/United States
- AG18440/AG/NIA NIH HHS/United States
- P01 AG022074/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
