

The replisome pausing factor Timeless is required for episomal maintenance of latent Epstein-Barr virus
- PMID: 21490103
- PMCID: PMC3126297
- DOI: 10.1128/JVI.02425-10
The replisome pausing factor Timeless is required for episomal maintenance of latent Epstein-Barr virus
Abstract
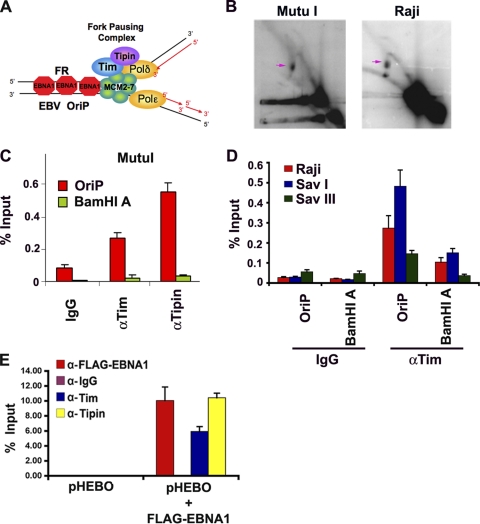
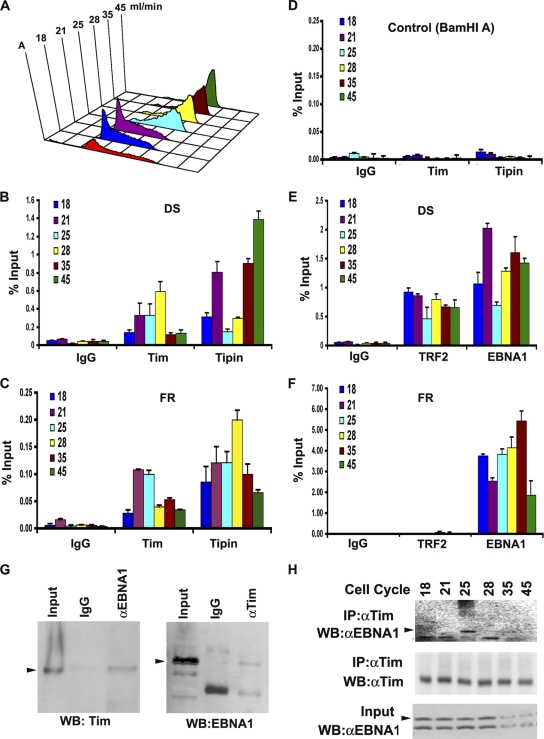
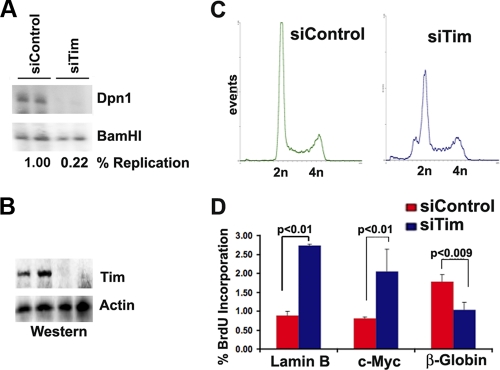
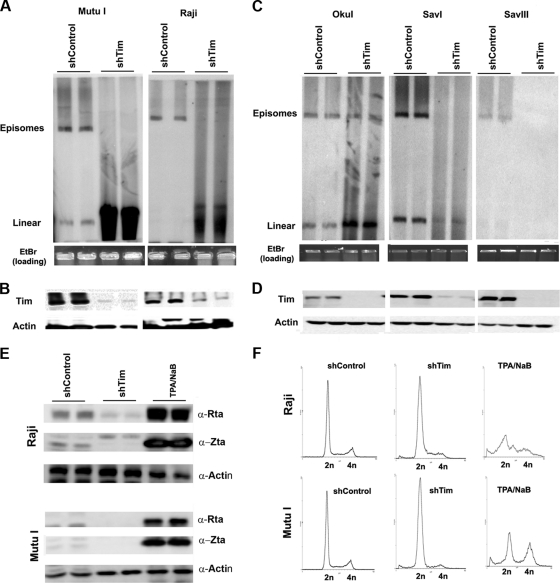
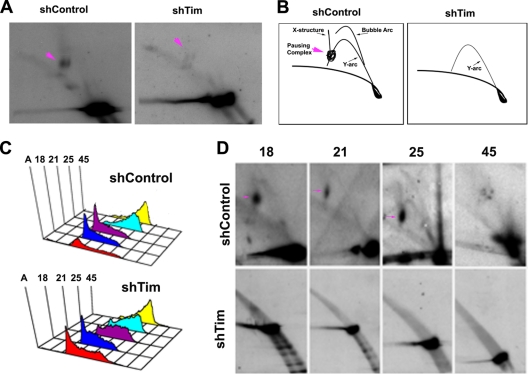
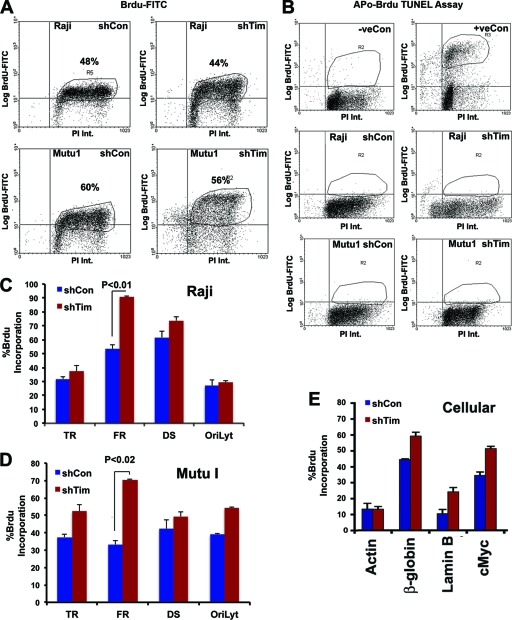
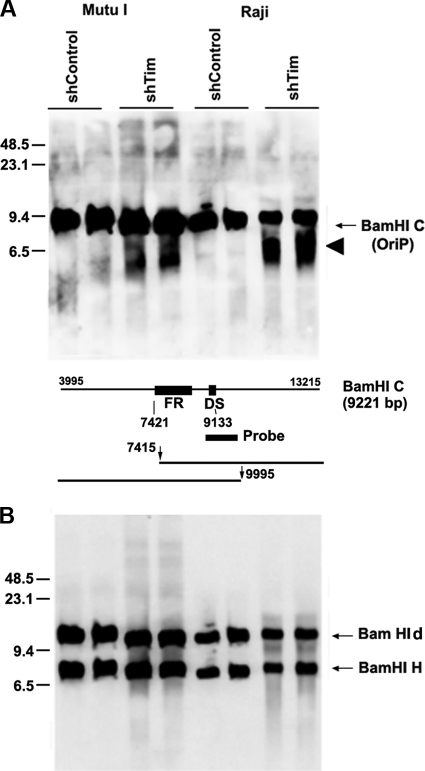
The Epstein-Barr virus (EBV) genome is maintained as an extrachromosomal episome during latent infection of B lymphocytes. Episomal maintenance is conferred by the interaction of the EBV-encoded nuclear antigen 1 (EBNA1) with a tandem array of high-affinity binding sites, referred to as the family of repeats (FR), located within the viral origin of plasmid replication (OriP). How this nucleoprotein array confers episomal maintenance is not completely understood. Previous studies have shown that DNA replication forks pause and terminate with high frequency at OriP. We now show that cellular DNA replication fork pausing and protection factors Timeless (Tim) and Tipin (Timeless-interacting protein) accumulate at OriP during S phase of the cell cycle. Depletion of Tim inhibits OriP-dependent DNA replication and causes a complete loss of the closed-circular form of EBV episomes in latently infected B lymphocytes. Tim depletion also led to the accumulation of double-strand breaks at the OriP region. These findings demonstrate that Tim is essential for sustaining the episomal forms of EBV DNA in latently infected cells and suggest that DNA replication fork protection is integrally linked to the mechanism of plasmid maintenance.
Figures
Similar articles
-
Timeless-dependent DNA replication-coupled recombination promotes Kaposi's Sarcoma-associated herpesvirus episome maintenance and terminal repeat stability.J Virol. 2013 Apr;87(7):3699-709. doi: 10.1128/JVI.02211-12. Epub 2013 Jan 16. J Virol. 2013. PMID: 23325691 Free PMC article.
-
Structural Basis for Cooperative Binding of EBNA1 to the Epstein-Barr Virus Dyad Symmetry Minimal Origin of Replication.J Virol. 2019 Sep 30;93(20):e00487-19. doi: 10.1128/JVI.00487-19. Print 2019 Oct 15. J Virol. 2019. PMID: 31142669 Free PMC article.
-
Cryo-EM Structure and Functional Studies of EBNA1 Binding to the Family of Repeats and Dyad Symmetry Elements of Epstein-Barr Virus oriP.J Virol. 2022 Sep 14;96(17):e0094922. doi: 10.1128/jvi.00949-22. Epub 2022 Aug 29. J Virol. 2022. PMID: 36037477 Free PMC article.
-
Replication licensing of the EBV oriP minichromosome.Curr Top Microbiol Immunol. 2001;258:13-33. doi: 10.1007/978-3-642-56515-1_2. Curr Top Microbiol Immunol. 2001. PMID: 11443858 Review.
-
[Epstein-Barr Virus Genome Replication as a Molecular Target for Cancer Therapy].Yakugaku Zasshi. 2019;139(1):63-67. doi: 10.1248/yakushi.18-00164-1. Yakugaku Zasshi. 2019. PMID: 30606931 Review. Japanese.
Cited by
-
Regulation of Telomere Homeostasis during Epstein-Barr virus Infection and Immortalization.Viruses. 2017 Aug 9;9(8):217. doi: 10.3390/v9080217. Viruses. 2017. PMID: 28792435 Free PMC article. Review.
-
Keeping it quiet: chromatin control of gammaherpesvirus latency.Nat Rev Microbiol. 2013 Dec;11(12):863-75. doi: 10.1038/nrmicro3135. Epub 2013 Nov 6. Nat Rev Microbiol. 2013. PMID: 24192651 Free PMC article. Review.
-
Timeless-dependent DNA replication-coupled recombination promotes Kaposi's Sarcoma-associated herpesvirus episome maintenance and terminal repeat stability.J Virol. 2013 Apr;87(7):3699-709. doi: 10.1128/JVI.02211-12. Epub 2013 Jan 16. J Virol. 2013. PMID: 23325691 Free PMC article.
-
An atlas of the Epstein-Barr virus transcriptome and epigenome reveals host-virus regulatory interactions.Cell Host Microbe. 2012 Aug 16;12(2):233-45. doi: 10.1016/j.chom.2012.06.008. Cell Host Microbe. 2012. PMID: 22901543 Free PMC article.
-
Separation of intra-S checkpoint protein contributions to DNA replication fork protection and genomic stability in normal human fibroblasts.Cell Cycle. 2013 Jan 15;12(2):332-45. doi: 10.4161/cc.23177. Epub 2012 Jan 15. Cell Cycle. 2013. PMID: 23255133 Free PMC article.
References
-
- Ballestas M. E., Chatis P. A., Kaye K. M. 1999. Efficient persistence of extrachromosomal KSHV DNA mediated by latency-associated nuclear antigen. Science 284:641–644 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
