

The stimulatory Gα(s) protein is involved in olfactory signal transduction in Drosophila
- PMID: 21490930
- PMCID: PMC3072409
- DOI: 10.1371/journal.pone.0018605
The stimulatory Gα(s) protein is involved in olfactory signal transduction in Drosophila
Abstract
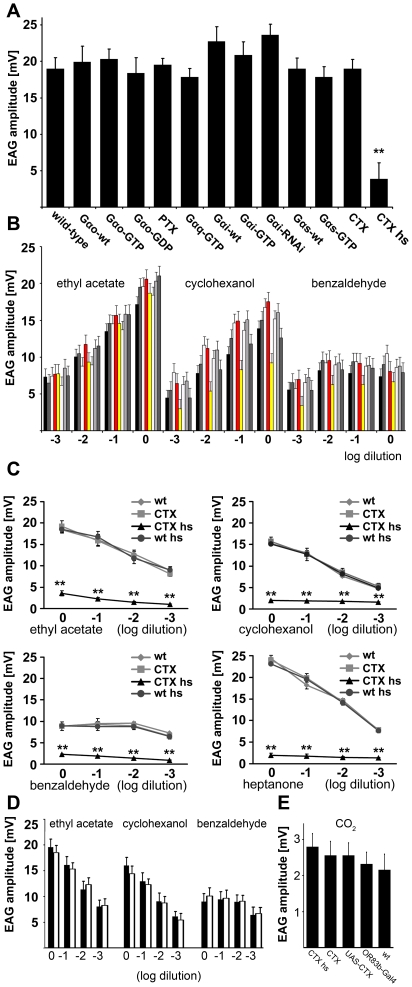
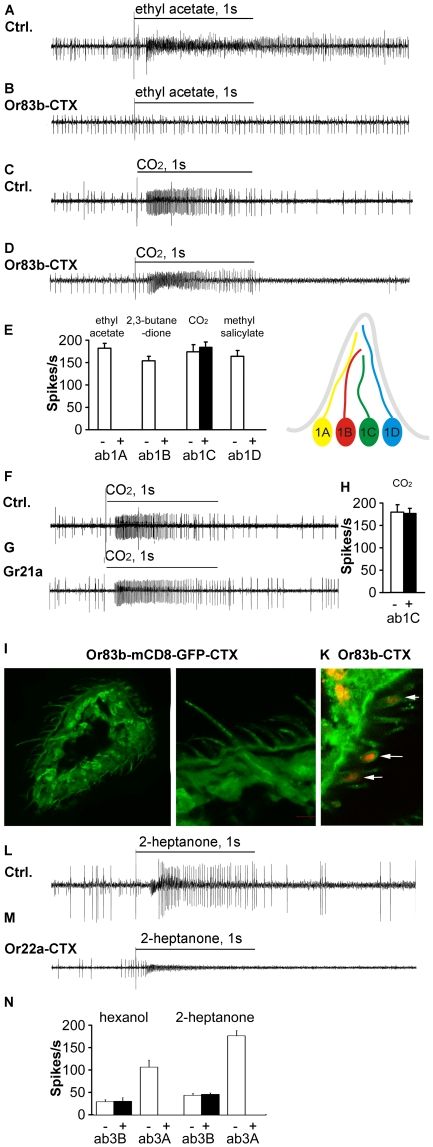
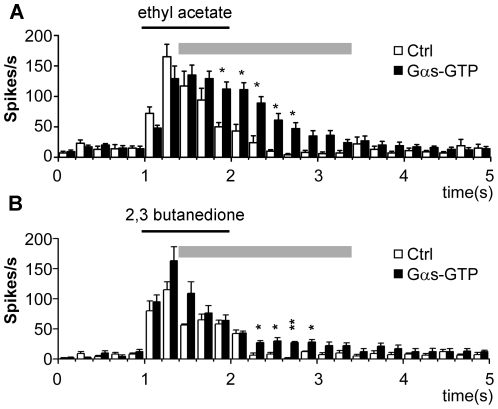
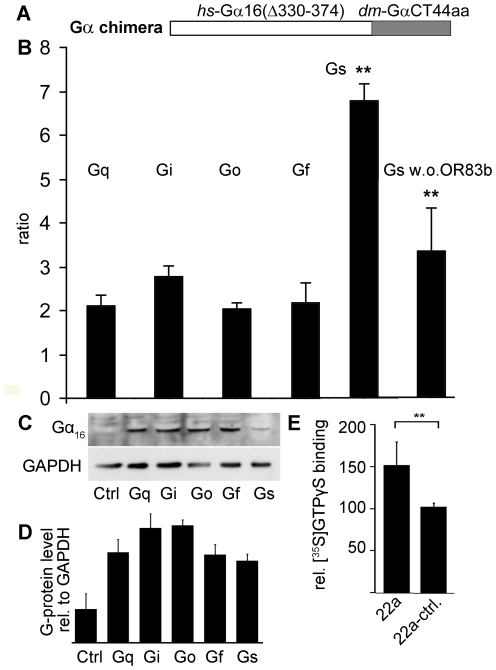
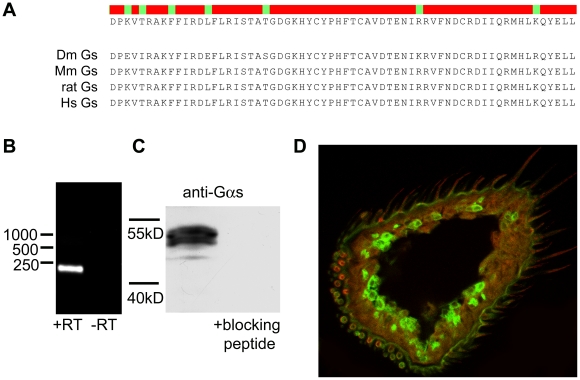
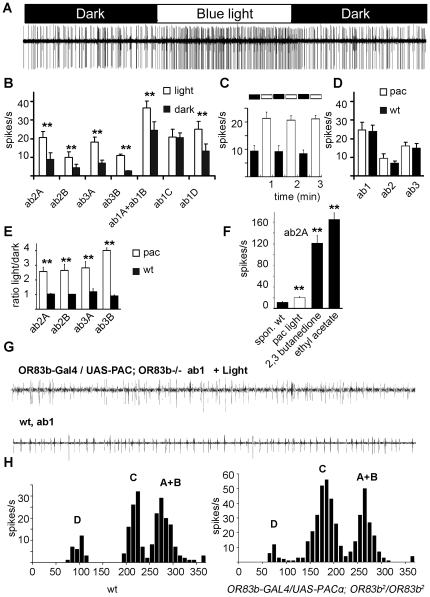
Seven-transmembrane receptors typically mediate olfactory signal transduction by coupling to G-proteins. Although insect odorant receptors have seven transmembrane domains like G-protein coupled receptors, they have an inverted membrane topology, constituting a key difference between the olfactory systems of insects and other animals. While heteromeric insect ORs form ligand-activated non-selective cation channels in recombinant expression systems, the evidence for an involvement of cyclic nucleotides and G-proteins in odor reception is inconsistent. We addressed this question in vivo by analyzing the role of G-proteins in olfactory signaling using electrophysiological recordings. We found that Gα(s) plays a crucial role for odorant induced signal transduction in OR83b expressing olfactory sensory neurons, but not in neurons expressing CO₂ responsive proteins GR21a/GR63a. Moreover, signaling of Drosophila ORs involved Gα(s) also in a heterologous expression system. In agreement with these observations was the finding that elevated levels of cAMP result in increased firing rates, demonstrating the existence of a cAMP dependent excitatory signaling pathway in the sensory neurons. Together, we provide evidence that Gα(s) plays a role in the OR mediated signaling cascade in Drosophila.
Conflict of interest statement
Figures
References
-
- Buck L, Axel R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 1991;65:175–187. - PubMed
-
- Firestein S. How the olfactory system makes sense of scents. Nature. 2001;413:211–218. - PubMed
-
- Ronnett GV, Moon C. G proteins and olfactory signal transduction. Annu Rev Physiol. 2002;64:189–222. - PubMed
-
- Vosshall LB, Wong AM, Axel R. An olfactory sensory map in the fly brain. Cell. 2000;102:147–159. - PubMed
-
- Nei M, Niimura Y, Nozawa M. The evolution of animal chemosensory receptor gene repertoires: roles of chance and necessity. Nat Rev Genet. 2008;9:951–963. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
