

Muscle pathology without severe nerve pathology in a new mouse model of Charcot-Marie-Tooth disease type 2E
- PMID: 21493625
- PMCID: PMC3109999
- DOI: 10.1093/hmg/ddr152
Muscle pathology without severe nerve pathology in a new mouse model of Charcot-Marie-Tooth disease type 2E
Abstract
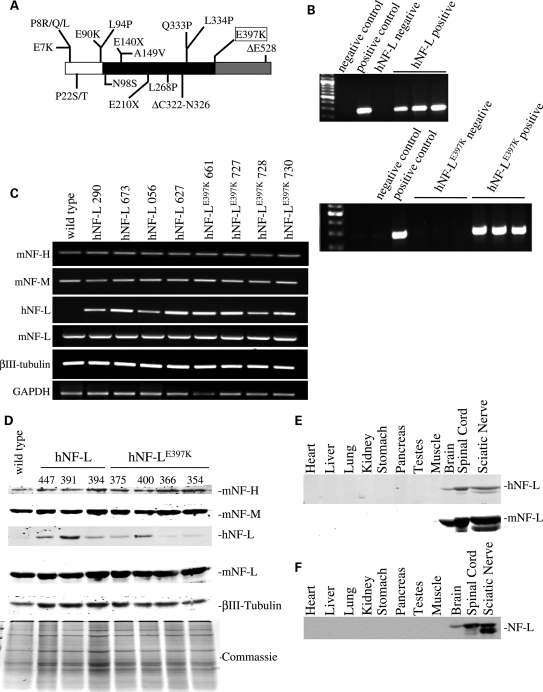
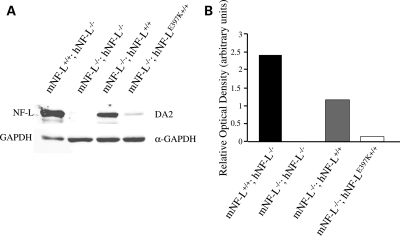
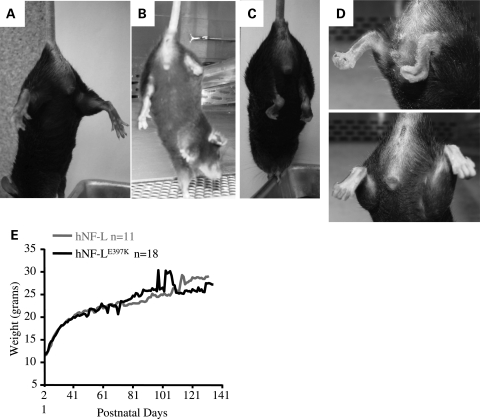
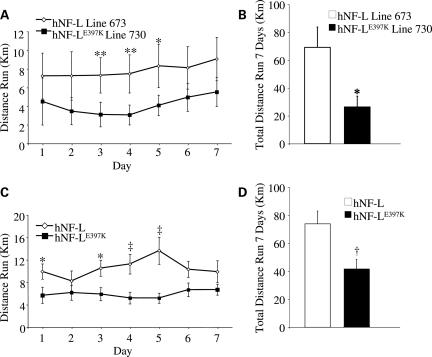
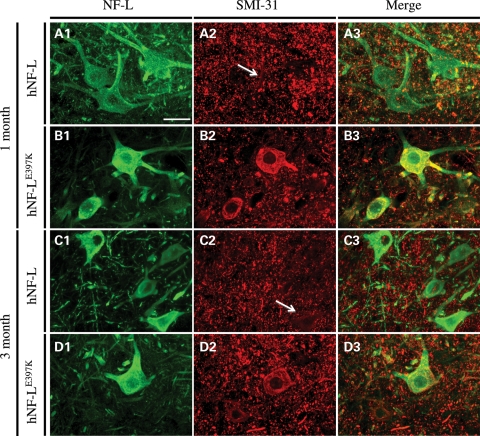
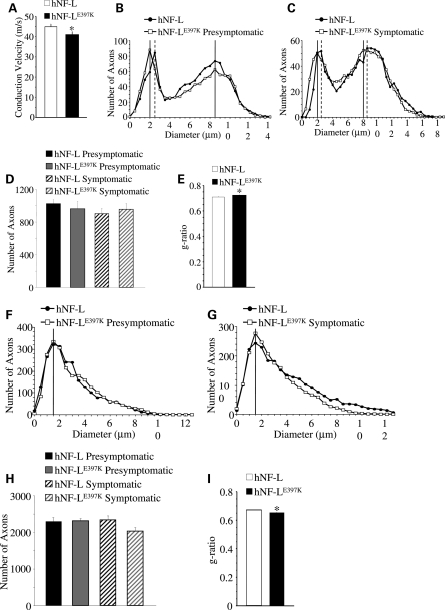
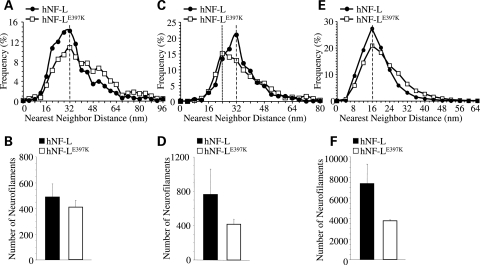
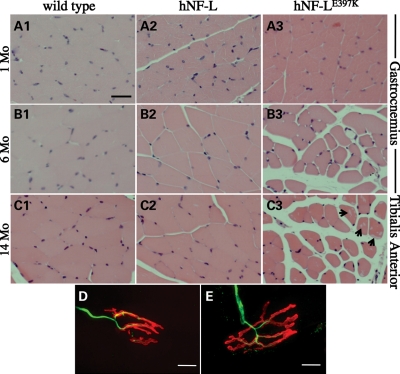
Mutations in neurofilament light (NF-L) have been linked to Charcot-Marie-Tooth disease type 2E (CMT2E) in humans. To provide insight into disease pathogenesis, we developed a novel line of CMT2E mice that constitutively express human NF-L (hNF-L) with a glutamic acid to lysine mutation at position 397 (hNF-L(E397K)). This new line of mice developed signs consistent with CMT2E patients. Disease signs were first observed at 4 months in hNF-L(E397K) mice, and consisted of aberrant hind limb posture, digit deformities, reduced voluntary locomotor activity, reduced motor nerve conduction velocities (MNCVs) and muscle atrophy. Reduced voluntary locomotor activity and muscle pathology occurred without significant denervation, and hNF-L(E397K) mice showed relatively mild signs of nerve pathology. Nerve pathology in hNF-L(E397K) mice was characterized by ectopic accumulations of phosphorylated NFs in motor neuron cell bodies as early as 1 month. Moreover, NF organization was altered in motor and sensory roots, with small motor axons being most affected. Peak axonal diameter was reduced for small motor axons prior to and after the onset of overt phenotypes, whereas large motor axons were affected only after onset, which correlated with reduced MNCVs. Additionally, there was a small reduction in the number of sensory axons in symptomatic hNF-L(E397K) mice. hNF-L(E397K) mice are a novel line of CMT2E mice that recapitulate many of the overt phenotypes observed in CMT2E patients and hNF-L(P22S) mice. The cellular pathology observed in hNF-L(E397K) mice differed from that recently reported in hNF-L(P22S) mice, suggesting that overt CMT2E phenotypes may arise through different cellular mechanisms.
Figures
Similar articles
-
Muscle spindle alterations precede onset of sensorimotor deficits in Charcot-Marie-Tooth type 2E.Genes Brain Behav. 2017 Feb;16(2):260-270. doi: 10.1111/gbb.12341. Epub 2016 Oct 11. Genes Brain Behav. 2017. PMID: 27643807 Free PMC article.
-
Expressing hNF-LE397K results in abnormal gaiting in a transgenic model of CMT2E.Genes Brain Behav. 2012 Apr;11(3):360-5. doi: 10.1111/j.1601-183X.2012.00771.x. Epub 2012 Feb 23. Genes Brain Behav. 2012. PMID: 22288874 Free PMC article.
-
Exacerbation of Charcot-Marie-Tooth type 2E neuropathy following traumatic nerve injury.Brain Res. 2015 Nov 19;1627:143-53. doi: 10.1016/j.brainres.2015.09.024. Epub 2015 Sep 28. Brain Res. 2015. PMID: 26423936 Free PMC article.
-
Demyelinating and axonal features of Charcot-Marie-Tooth disease with mutations of myelin-related proteins (PMP22, MPZ and Cx32): a clinicopathological study of 205 Japanese patients.Brain. 2003 Jan;126(Pt 1):134-51. doi: 10.1093/brain/awg012. Brain. 2003. PMID: 12477701 Review.
-
Axonal Charcot-Marie-Tooth disease.Curr Opin Neurol. 2011 Oct;24(5):475-83. doi: 10.1097/WCO.0b013e32834aa331. Curr Opin Neurol. 2011. PMID: 21892080 Review.
Cited by
-
Neurofilament light polypeptide gene N98S mutation in mice leads to neurofilament network abnormalities and a Charcot-Marie-Tooth Type 2E phenotype.Hum Mol Genet. 2015 Apr 15;24(8):2163-74. doi: 10.1093/hmg/ddu736. Epub 2014 Dec 30. Hum Mol Genet. 2015. PMID: 25552649 Free PMC article.
-
Muscle spindle alterations precede onset of sensorimotor deficits in Charcot-Marie-Tooth type 2E.Genes Brain Behav. 2017 Feb;16(2):260-270. doi: 10.1111/gbb.12341. Epub 2016 Oct 11. Genes Brain Behav. 2017. PMID: 27643807 Free PMC article.
-
Dental pathologies in lamniform and carcharhiniform sharks with comments on the classification and homology of double tooth pathologies in vertebrates.PeerJ. 2022 May 11;10:e12775. doi: 10.7717/peerj.12775. eCollection 2022. PeerJ. 2022. PMID: 35578672 Free PMC article.
-
Neurofilament Phosphorylation during Development and Disease: Which Came First, the Phosphorylation or the Accumulation?J Amino Acids. 2012;2012:382107. doi: 10.1155/2012/382107. Epub 2012 Apr 18. J Amino Acids. 2012. PMID: 22570767 Free PMC article.
-
Neurofilament Proteins as Biomarkers to Monitor Neurological Diseases and the Efficacy of Therapies.Front Neurosci. 2021 Sep 27;15:689938. doi: 10.3389/fnins.2021.689938. eCollection 2021. Front Neurosci. 2021. PMID: 34646114 Free PMC article. Review.
References
-
- Skre H. Genetic and clinical aspects of Charcot–Marie–Tooth's disease. Clin. Genet. 1974;6:98–118. doi:10.1111/j.1399-0004.1974.tb00638.x. - DOI - PubMed
-
- Emery A.E. Population frequencies of inherited neuromuscular diseases—a world survey. Neuromuscul. Disord. 1991;1:19–29. doi:10.1016/0960-8966(91)90039-U. - DOI - PubMed
-
- Mersiyanova I.V., Perepelov A.V., Polyakov A.V., Sitnikov V.F., Dadali E.L., Oparin R.B., Petrin A.N., Evgrafov O.V. A new variant of Charcot–Marie–Tooth disease type 2 is probably the result of a mutation in the neurofilament-light gene. Am. J. Hum. Genet. 2000;67:37–46. doi:10.1086/302962. - DOI - PMC - PubMed
-
- De Jonghe P., Mersivanova I., Nelis E., Del Favero J., Martin J.J., Van Broeckhoven C., Evgrafov O., Timmerman V. Further evidence that neurofilament light chain gene mutations can cause Charcot–Marie–Tooth disease type 2E. Ann. Neurol. 2001;49:245–249. doi:10.1002/1531-8249(20010201)49:2<245::AID-ANA45>3.0.CO;2-A. - DOI - PubMed
-
- Georgiou D.M., Zidar J., Korosec M., Middleton L.T., Kyriakides T., Christodoulou K. A novel NF-L mutation Pro22Ser is associated with CMT2 in a large Slovenian family. Neurogenetics. 2002;4:93–96. doi:10.1007/s10048-002-0138-4. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
