

A 5'-proximal stem-loop structure of 5' untranslated region of porcine reproductive and respiratory syndrome virus genome is key for virus replication
- PMID: 21496223
- PMCID: PMC3096946
- DOI: 10.1186/1743-422X-8-172
A 5'-proximal stem-loop structure of 5' untranslated region of porcine reproductive and respiratory syndrome virus genome is key for virus replication
Abstract
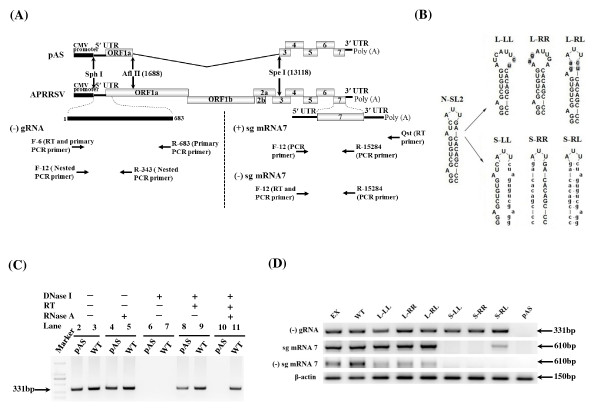
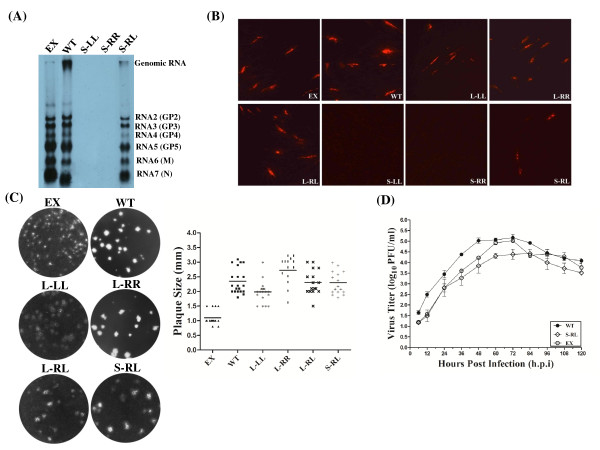
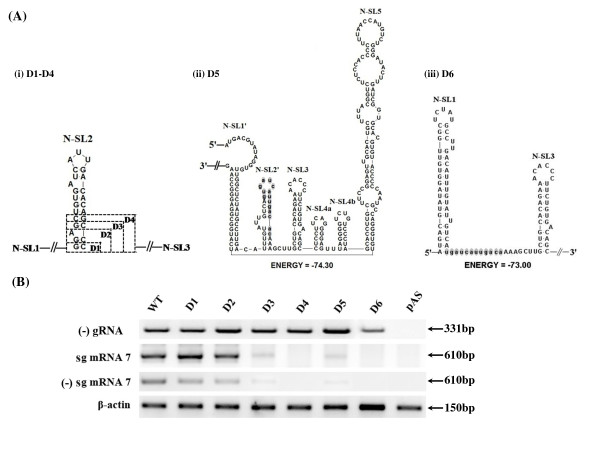
Background: It has been well documented that the 5' untranslated region (5' UTR) of many positive-stranded RNA viruses contain key cis-acting regulatory sequences, as well as high-order structural elements. Little is known for such regulatory elements controlling porcine arterivirus replication. We investigated the roles of a conserved stem-loop 2 (SL2) that resides in the 5'UTR of the genome of a type II porcine reproductive and respiratory syndrome virus (PRRSV).
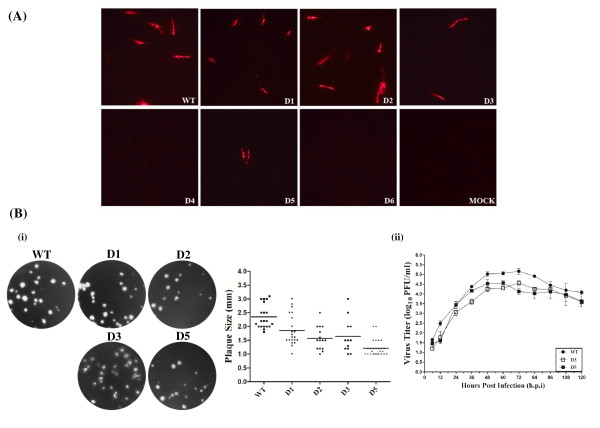
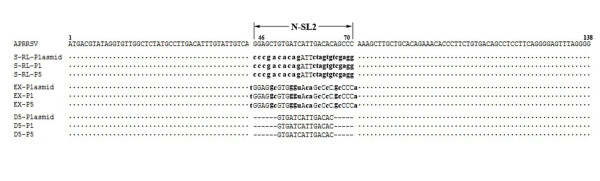
Results: We provided genetic evidences demonstrating that 1) the SL2 in type II PRRSV 5' UTR, N-SL2, could be structurally and functionally substituted by its counterpart in type I PRRSV, E-SL2; 2) the functionality of N-SL2 was dependent upon the G-C rich stem structure, while the ternary-loop size was irrelevant to RNA synthesis; 3) serial deletions showed that the stem integrity of N-SL2 was crucial for subgenomic mRNA synthesis; and 4) when extensive base-pairs in the stem region was deleted, an alternative N-SL2-like structure with different sequence was utilized for virus replication.
Conclusion: Taken together, we concluded that the phylogenetically conserved SL2 in the 5' UTR was crucial for PRRSV virus replication, subgenomic mRNA synthesis in particular.
Figures
Similar articles
-
Replacement of the heterologous 5' untranslated region allows preservation of the fully functional activities of type 2 porcine reproductive and respiratory syndrome virus.Virology. 2013 Apr 25;439(1):1-12. doi: 10.1016/j.virol.2012.12.013. Epub 2013 Mar 1. Virology. 2013. PMID: 23453581 Free PMC article.
-
Cis-acting structural element in 5' UTR is essential for infectivity of porcine reproductive and respiratory syndrome virus.Virus Res. 2012 Jan;163(1):108-19. doi: 10.1016/j.virusres.2011.08.018. Epub 2011 Sep 8. Virus Res. 2012. PMID: 21924304 Free PMC article.
-
Identification of dispensable nucleotide sequence in 3' untranslated region of porcine reproductive and respiratory syndrome virus.Virus Res. 2010 Dec;154(1-2):38-47. doi: 10.1016/j.virusres.2010.08.027. Epub 2010 Sep 15. Virus Res. 2010. PMID: 20833212 Free PMC article. Review.
-
Putative cis-acting stem-loops in the 5' untranslated region of the severe acute respiratory syndrome coronavirus can substitute for their mouse hepatitis virus counterparts.J Virol. 2006 Nov;80(21):10600-14. doi: 10.1128/JVI.00455-06. Epub 2006 Aug 18. J Virol. 2006. PMID: 16920822 Free PMC article.
-
PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity.Virology. 2015 May;479-480:475-86. doi: 10.1016/j.virol.2015.02.012. Epub 2015 Mar 7. Virology. 2015. PMID: 25759097 Free PMC article. Review.
Cited by
-
Genomic sequencing reveals mutations potentially related to the overattenuation of a highly pathogenic porcine reproductive and respiratory syndrome virus.Clin Vaccine Immunol. 2013 Apr;20(4):613-9. doi: 10.1128/CVI.00672-12. Epub 2013 Feb 13. Clin Vaccine Immunol. 2013. PMID: 23408525 Free PMC article.
-
The Nsp12-coding region of type 2 PRRSV is required for viral subgenomic mRNA synthesis.Emerg Microbes Infect. 2019;8(1):1501-1510. doi: 10.1080/22221751.2019.1679010. Emerg Microbes Infect. 2019. PMID: 31631782 Free PMC article.
-
In-Depth Bioinformatic Analyses of Nidovirales Including Human SARS-CoV-2, SARS-CoV, MERS-CoV Viruses Suggest Important Roles of Non-canonical Nucleic Acid Structures in Their Lifecycles.Front Microbiol. 2020 Jul 3;11:1583. doi: 10.3389/fmicb.2020.01583. eCollection 2020. Front Microbiol. 2020. PMID: 32719673 Free PMC article.
-
Replacement of the heterologous 5' untranslated region allows preservation of the fully functional activities of type 2 porcine reproductive and respiratory syndrome virus.Virology. 2013 Apr 25;439(1):1-12. doi: 10.1016/j.virol.2012.12.013. Epub 2013 Mar 1. Virology. 2013. PMID: 23453581 Free PMC article.
-
Structural Characterization of Non-structural Protein 9 Complexed With Specific Nanobody Pinpoints Two Important Residues Involved in Porcine Reproductive and Respiratory Syndrome Virus Replication.Front Microbiol. 2020 Nov 12;11:581856. doi: 10.3389/fmicb.2020.581856. eCollection 2020. Front Microbiol. 2020. PMID: 33281776 Free PMC article.
References
-
- Snijder EJ, Meulenberg JJ. The molecular biology of arteriviruses. J Gen Virol. 1998;79(Pt 5):961–979. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
