

Caveolin-1 deficiency causes cholesterol-dependent mitochondrial dysfunction and apoptotic susceptibility
- PMID: 21497090
- PMCID: PMC3409647
- DOI: 10.1016/j.cub.2011.03.030
Caveolin-1 deficiency causes cholesterol-dependent mitochondrial dysfunction and apoptotic susceptibility
Abstract
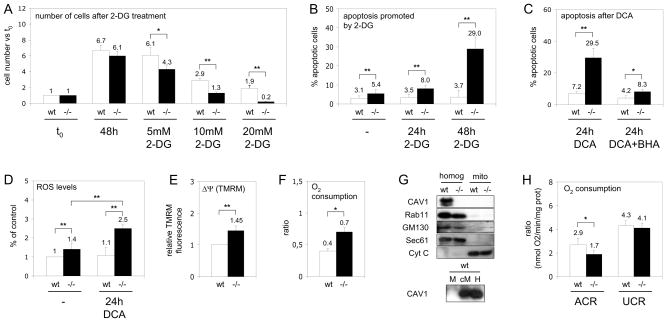
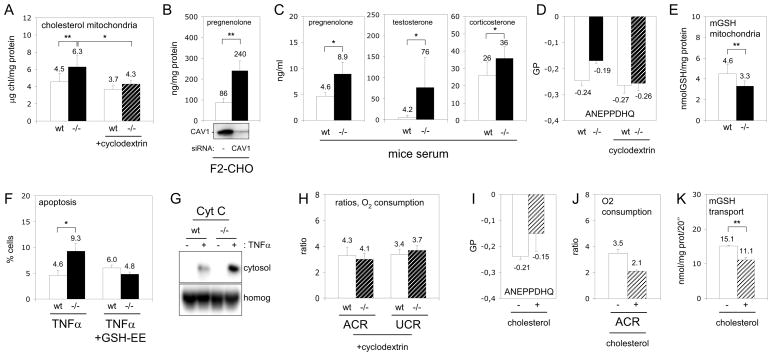
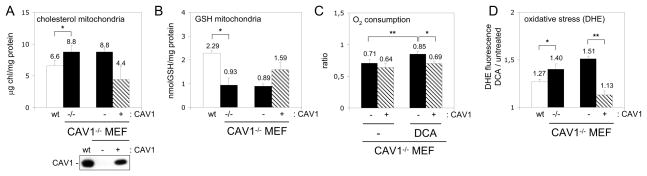
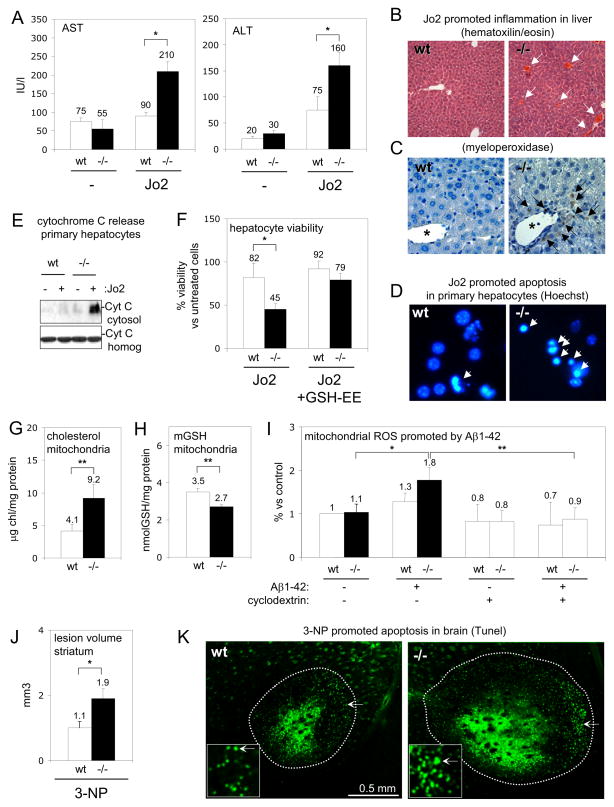
Caveolins (CAVs) are essential components of caveolae, plasma membrane invaginations with reduced fluidity, reflecting cholesterol accumulation. CAV proteins bind cholesterol, and CAV's ability to move between cellular compartments helps control intracellular cholesterol fluxes. In humans, CAV1 mutations result in lipodystrophy, cell transformation, and cancer. CAV1 gene-disrupted mice exhibit cardiovascular diseases, diabetes, cancer, atherosclerosis, and pulmonary fibrosis. The mechanism or mechanisms underlying these disparate effects are unknown, but our past work suggested that CAV1 deficiency might alter metabolism: CAV1(-/-) mice exhibit impaired liver regeneration unless supplemented with glucose, suggesting systemic inefficiencies requiring additional metabolic intermediates. Establishing a functional link between CAV1 and metabolism would provide a unifying theme to explain these myriad pathologies. Here we demonstrate that impaired proliferation and low survival with glucose restriction is a shortcoming of CAV1-deficient cells caused by impaired mitochondrial function. Without CAV1, free cholesterol accumulates in mitochondrial membranes, increasing membrane condensation and reducing efficiency of the respiratory chain and intrinsic antioxidant defense. Upon activation of oxidative phosphorylation, this promotes accumulation of reactive oxygen species, resulting in cell death. We confirm that this mitochondrial dysfunction predisposes CAV1-deficient animals to mitochondrial-related diseases such as steatohepatitis and neurodegeneration.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Parton RG, Simons K. The multiple faces of caveolae. Nat Rev Mol Cell Biol. 2007;8:185–194. - PubMed
-
- Kim CA, Delepine M, Boutet E, El Mourabit H, Le Lay S, Meier M, Nemani M, Bridel E, Leite CC, Bertola DR, et al. Association of a homozygous nonsense caveolin-1 mutation with Berardinelli-Seip congenital lipodystrophy. J Clin Endocrinol Metab. 2008;93:1129–1134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
