

Competing interactions stabilize pro- and anti-aggregant conformations of human Tau
- PMID: 21498513
- PMCID: PMC3121454
- DOI: 10.1074/jbc.M111.237875
Competing interactions stabilize pro- and anti-aggregant conformations of human Tau
Abstract
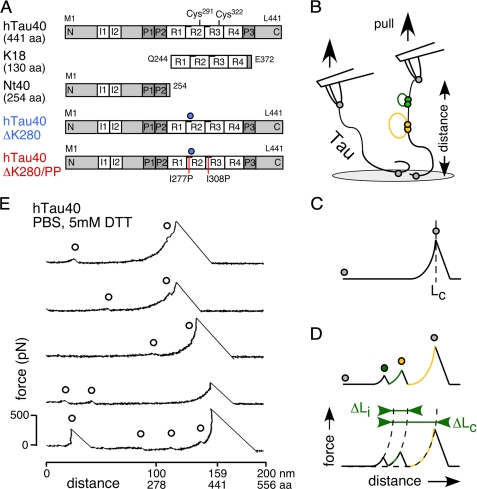
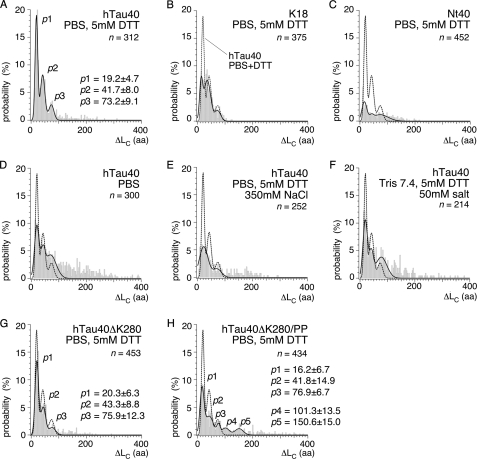
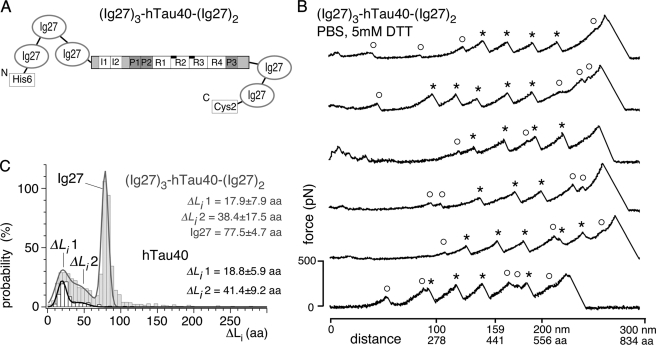
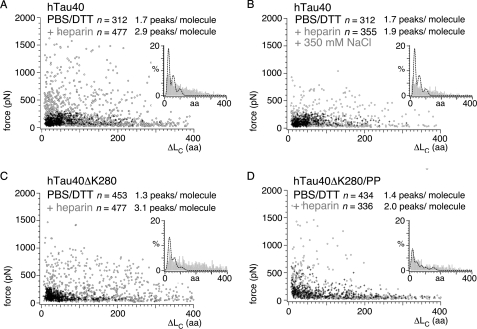
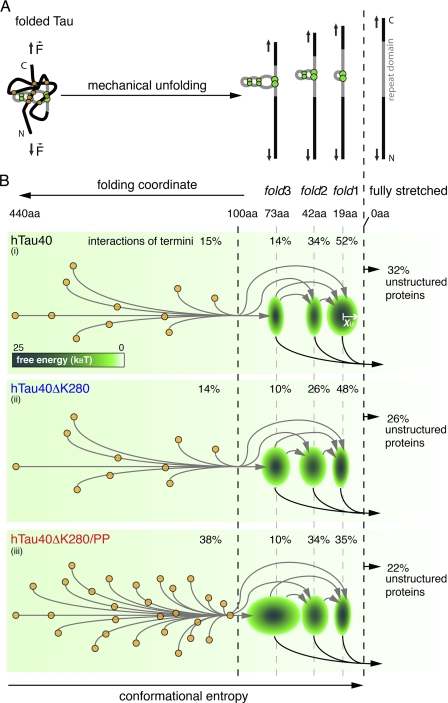
Aggregation of Tau into amyloid-like fibrils is a key process in neurodegenerative diseases such as Alzheimer. To understand how natively disordered Tau stabilizes conformations that favor pathological aggregation, we applied single-molecule force spectroscopy. Intramolecular interactions that fold polypeptide stretches of ~19 and ~42 amino acids in the functionally important repeat domain of full-length human Tau (hTau40) support aggregation. In contrast, the unstructured N terminus randomly folds long polypeptide stretches >100 amino acids that prevent aggregation. The pro-aggregant mutant hTau40ΔK280 observed in frontotemporal dementia favored the folding of short polypeptide stretches and suppressed the folding of long ones. This trend was reversed in the anti-aggregant mutant hTau40ΔK280/PP. The aggregation inducer heparin introduced strong interactions in hTau40 and hTau40ΔK280 that stabilized aggregation-prone conformations. We show that the conformation and aggregation of Tau are regulated through a complex balance of different intra- and intermolecular interactions.
Figures
Similar articles
-
Regulatable transgenic mouse models of Alzheimer disease: onset, reversibility and spreading of Tau pathology.FEBS J. 2013 Sep;280(18):4371-81. doi: 10.1111/febs.12250. Epub 2013 Apr 22. FEBS J. 2013. PMID: 23517246 Review.
-
Stages and conformations of the Tau repeat domain during aggregation and its effect on neuronal toxicity.J Biol Chem. 2014 Jul 18;289(29):20318-32. doi: 10.1074/jbc.M114.554725. Epub 2014 May 13. J Biol Chem. 2014. PMID: 24825901 Free PMC article.
-
Conformational diversity of wild-type Tau fibrils specified by templated conformation change.J Biol Chem. 2009 Feb 6;284(6):3546-51. doi: 10.1074/jbc.M805627200. Epub 2008 Nov 14. J Biol Chem. 2009. PMID: 19010781 Free PMC article.
-
Spreading of Tau Protein Does Not Depend on Aggregation Propensity.J Mol Neurosci. 2023 Oct;73(9-10):693-712. doi: 10.1007/s12031-023-02143-w. Epub 2023 Aug 22. J Mol Neurosci. 2023. PMID: 37606769 Free PMC article.
-
Amyloidogenesis of Tau protein.Protein Sci. 2017 Nov;26(11):2126-2150. doi: 10.1002/pro.3275. Epub 2017 Sep 13. Protein Sci. 2017. PMID: 28833749 Free PMC article. Review.
Cited by
-
Alternative conformations of the Tau repeat domain in complex with an engineered binding protein.J Biol Chem. 2014 Aug 15;289(33):23209-23218. doi: 10.1074/jbc.M114.560920. Epub 2014 Jun 25. J Biol Chem. 2014. PMID: 24966331 Free PMC article.
-
Mechanical Effects of Dynamic Binding between Tau Proteins on Microtubules during Axonal Injury.Biophys J. 2015 Dec 1;109(11):2328-37. doi: 10.1016/j.bpj.2015.09.010. Biophys J. 2015. PMID: 26636944 Free PMC article.
-
Identification of an aggregation-prone structure of tau.J Am Chem Soc. 2012 Oct 10;134(40):16607-13. doi: 10.1021/ja305206m. Epub 2012 Oct 1. J Am Chem Soc. 2012. PMID: 22998648 Free PMC article.
-
Physical Biology of Axonal Damage.Front Cell Neurosci. 2018 Jun 6;12:144. doi: 10.3389/fncel.2018.00144. eCollection 2018. Front Cell Neurosci. 2018. PMID: 29928193 Free PMC article.
-
Taxol-stabilized microtubules promote the formation of filaments from unmodified full-length Tau in vitro.Mol Biol Cell. 2012 Dec;23(24):4796-806. doi: 10.1091/mbc.E12-05-0374. Epub 2012 Oct 19. Mol Biol Cell. 2012. PMID: 23087208 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
