

Review
doi: 10.1093/nar/gkr223.
Epub 2011 Apr 15.
How does DNA break during chromosomal translocations?
Affiliations
- PMID: 21498543
- PMCID: PMC3152359
- DOI: 10.1093/nar/gkr223
Item in Clipboard
Review
How does DNA break during chromosomal translocations?
Nucleic Acids Res.
2011 Aug.
Abstract
Chromosomal translocations are one of the most common types of genetic rearrangements and are molecular signatures for many types of cancers. They are considered as primary causes for cancers, especially lymphoma and leukemia. Although many translocations have been reported in the last four decades, the mechanism by which chromosomes break during a translocation remains largely unknown. In this review, we summarize recent advances made in understanding the molecular mechanism of chromosomal translocations.
Figures
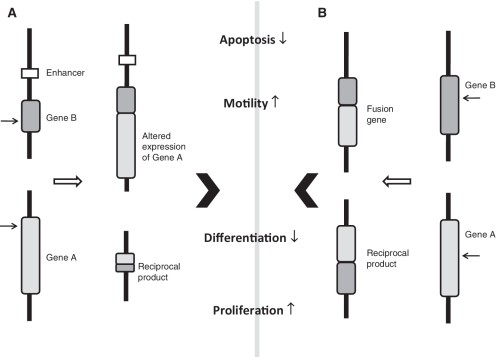
Consequences of chromosomal translocations. A chromosomal translocation can broadly result in either juxtaposition of oncogenes near promoter/enhancer elements (A) or gene fusions (B). Both results in the deregulation of the expression of genes affecting various cellular and physiological processes like proliferation, differentiation, motility and apoptosis. Arrows at the genes depict the sites for double-strand break formation. Normally, the reciprocal joining does not lead to any functional product.
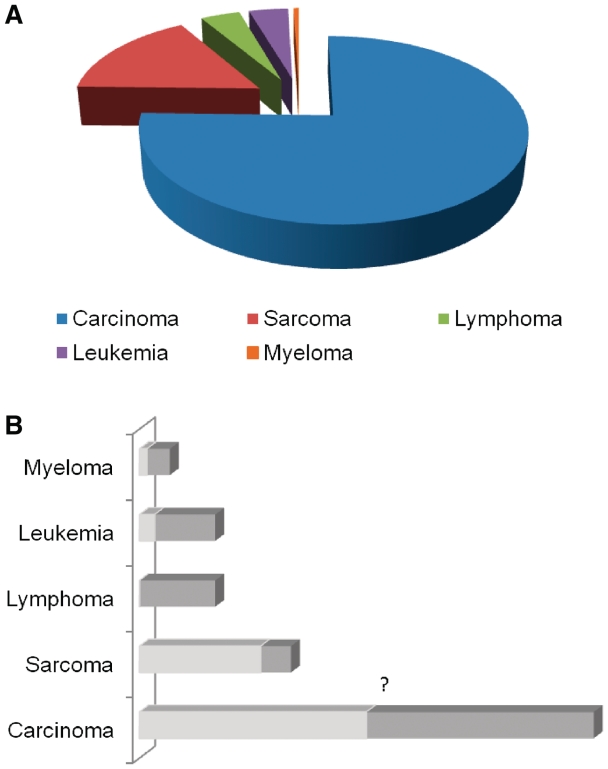
Frequency of chromosomal translocations in different cancers. (A) Distribution of different classes of cancers in terms of their incidence. Carcinoma constitutes the maximum percentage among all types of cancer followed by sarcoma. Leukemia and lymphoma together constitute around 8% of all cancers. (B) Percentage of occurrence of translocations within the different classes of cancer. Almost all lymphomas harbor translocations, whereas only one-fourth of all sarcomas (183) have been reported to possess the same. Gene fusions in carcinomas have been discovered recently. Although recent investigations suggest its presence in many carcinomas, the exact percentage of carcinoma bearing translocations is not yet clear (denoted by ‘?’).
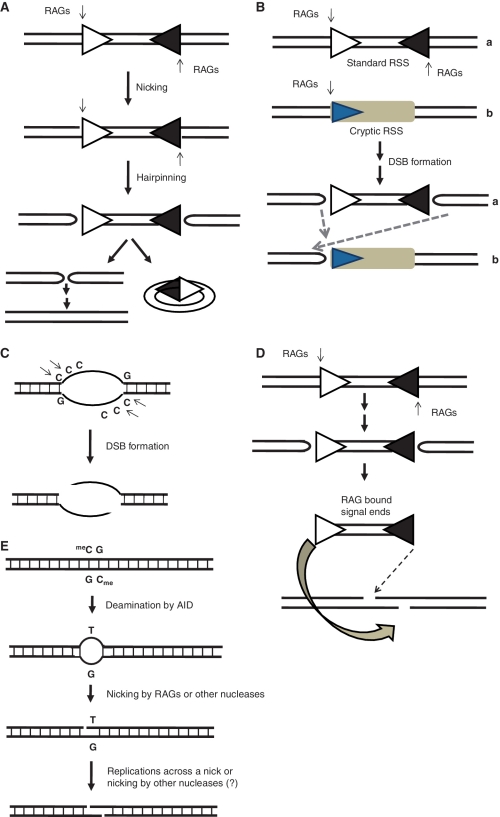
Mechanism of RAG mediated translocations. (A) Standard V(D)J recombination is initiated by RAG induced nicks (indicated by thin arrows) at the 5′ of the heptamer present within the standard recombination signal sequences (open triangle represents 12 RSS while closed triangle depicts 23 RSS). This is followed by hairpin formation using a transesterification reaction. The hairpins are further resolved and DSBs are joined using NHEJ protein machinery resulting in the formation of a coding joint. The free signal ends circularize to give the signal joint. (B) RAGs, in addition to nicking the standard recombination signals (a), can misrecognize sequences resembling standard RSS on other genes (b) and induce nicks, which can be converted to hairpins, using the standard V(D)J-like mechanism. These hairpins can then be processed and joined using the components of the NHEJ pathway. (C) RAGs can also recognize single-stranded regions of non-B DNA structures, especially in the context of cytosines and induce nicks or DSBs. (D) RAG-mediated transposition can also introduce genomic instability and hence translocations. RAGs can act like a transposase and introduce signal ends into heterologous chromosomes at some other loci making them fragile. (E) CpGs are susceptible to deamination by enzymes like AID, resulting in single-nucleotide mismatches. These can in turn be recognized and cleaved by RAGs or other nucleases, to introduce nicks or DSBs. Such CpGs have been found at or near many chromosomal translocation breakpoint regions in lymphoid cancers.
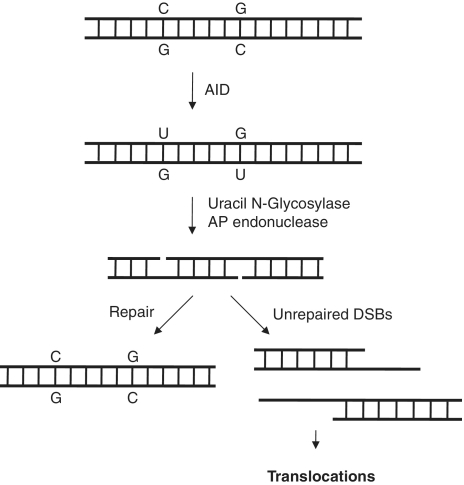
Mechanism of AID-mediated translocations. AID can deaminate cytosines leading to the formation of uracil, which is generally removed by Uracil N–Glycosylase. The nicks are usually repaired by the base excision repair mechanism. However, two unrepaired nicks in close vicinity can act as a double-strand break and thereby a substrate for chromosomal translocations.
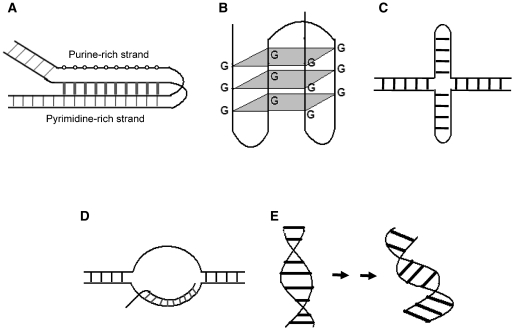
Non-B DNA structures involved in generation of chromosomal translocations. Recent studies have shown the importance of altered or non-B DNA structures in the generation of chromosomal translocations. Triplex DNA (A), G-quadruplexes (B), cruciform DNA (C), RNA-DNA hybrid (D) and B/A intermediate (E) DNA structures have been reported to be present at several ommon translocation breakpoint sites.
References
-
- Boveri T. Zur Frage der Entstehung maligner Tumoren [The Origin of Malignant Tumors] 1914. Gustav Fischer, Jena.
-
- Stern C. Boveri and the early days of genetics. Nature. 1950;166:446. - PubMed
-
- Nowell PC, Hungerford DA. A minute chromosome in human chronic granulocytic leukemia. Science. 1960;132:1497. - PubMed
-
- Rowley JD. A new consistent chromosomal abnormality in chronic Myelogenous leukaemia identified by quinacrine fluorescence and Giemsa staining. Nature. 1973;243:290–293. - PubMed
-
- Zech L, Haglund U, Nilsson K, Klein G. Characteristic chromosomal abnormalities in biopsies and lymphoid-cell lines from patients with Burkitt and non-Burkitt lymphomas. Int. J. Cancer. 1976;17:47–56. - PubMed
