

DYRK1A protein kinase promotes quiescence and senescence through DREAM complex assembly
- PMID: 21498570
- PMCID: PMC3078706
- DOI: 10.1101/gad.2034211
DYRK1A protein kinase promotes quiescence and senescence through DREAM complex assembly
Abstract
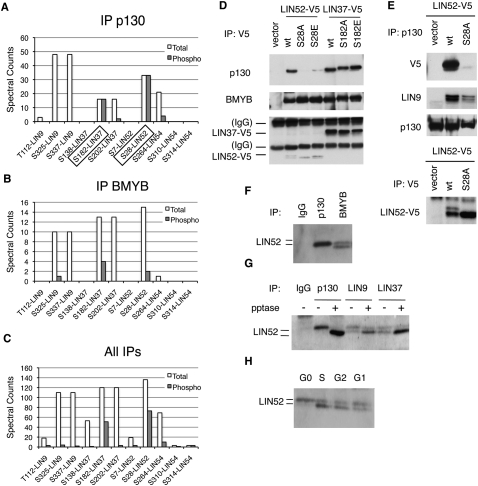
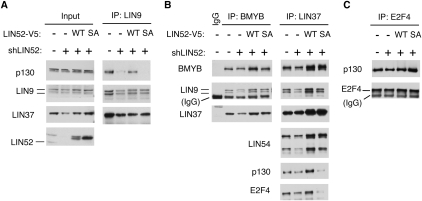
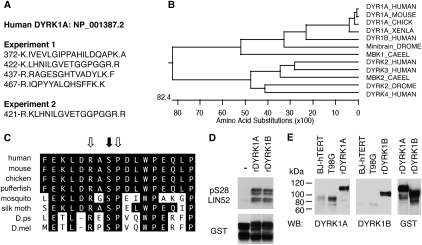
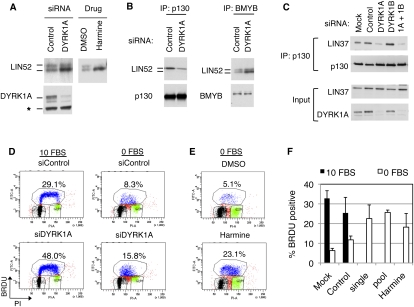
In the absence of growth signals, cells exit the cell cycle and enter into G0 or quiescence. Alternatively, cells enter senescence in response to inappropriate growth signals such as oncogene expression. The molecular mechanisms required for cell cycle exit into quiescence or senescence are poorly understood. The DREAM (DP, RB [retinoblastoma], E2F, and MuvB) complex represses cell cycle-dependent genes during quiescence. DREAM contains p130, E2F4, DP1, and a stable core complex of five MuvB-like proteins: LIN9, LIN37, LIN52, LIN54, and RBBP4. In mammalian cells, the MuvB core dissociates from p130 upon entry into the cell cycle and binds to BMYB during S phase to activate the transcription of genes expressed late in the cell cycle. We used mass spectroscopic analysis to identify phosphorylation sites that regulate the switch of the MuvB core from BMYB to DREAM. Here we report that DYRK1A can specifically phosphorylate LIN52 on serine residue 28, and that this phosphorylation is required for DREAM assembly. Inhibiting DYRK1A activity or point mutation of LIN52 disrupts DREAM assembly and reduces the ability of cells to enter quiescence or undergo Ras-induced senescence. These data reveal an important role for DYRK1A in the regulation of DREAM activity and entry into quiescence.
Figures
Comment in
-
Sweet DREAMs for Hippo.Genes Dev. 2011 May 1;25(9):889-94. doi: 10.1101/gad.2050411. Genes Dev. 2011. PMID: 21536729 Free PMC article.
Similar articles
-
Structural mechanisms of DREAM complex assembly and regulation.Genes Dev. 2015 May 1;29(9):961-74. doi: 10.1101/gad.257568.114. Epub 2015 Apr 27. Genes Dev. 2015. PMID: 25917549 Free PMC article.
-
Evolutionarily conserved multisubunit RBL2/p130 and E2F4 protein complex represses human cell cycle-dependent genes in quiescence.Mol Cell. 2007 May 25;26(4):539-51. doi: 10.1016/j.molcel.2007.04.015. Mol Cell. 2007. PMID: 17531812
-
Loss of the mammalian DREAM complex deregulates chondrocyte proliferation.Mol Cell Biol. 2014 Jun;34(12):2221-34. doi: 10.1128/MCB.01523-13. Epub 2014 Apr 7. Mol Cell Biol. 2014. PMID: 24710275 Free PMC article.
-
MuvB: A Key to Cell Cycle Control in Ovarian Cancer.Front Oncol. 2018 Jun 11;8:223. doi: 10.3389/fonc.2018.00223. eCollection 2018. Front Oncol. 2018. PMID: 29942794 Free PMC article. Review.
-
Activity of the retinoblastoma family proteins, pRB, p107, and p130, during cellular proliferation and differentiation.Crit Rev Biochem Mol Biol. 1996 Jun;31(3):237-71. doi: 10.3109/10409239609106585. Crit Rev Biochem Mol Biol. 1996. PMID: 8817077 Review.
Cited by
-
Tumorigenesis in Down's syndrome: big lessons from a small chromosome.Nat Rev Cancer. 2012 Oct;12(10):721-32. doi: 10.1038/nrc3355. Epub 2012 Sep 21. Nat Rev Cancer. 2012. PMID: 22996602
-
Differential regulation of expression of the protein kinases DYRK1A and DYRK1B in cancer cells.Sci Rep. 2024 Oct 13;14(1):23926. doi: 10.1038/s41598-024-74190-1. Sci Rep. 2024. PMID: 39397076 Free PMC article.
-
Stable Phenotypic Changes of the Host T Cells Are Essential to the Long-Term Stability of Latent HIV-1 Infection.J Virol. 2015 Jul;89(13):6656-72. doi: 10.1128/JVI.00571-15. Epub 2015 Apr 15. J Virol. 2015. PMID: 25878110 Free PMC article.
-
The biology, pathogenesis and clinical aspects of acute lymphoblastic leukemia in children with Down syndrome.Leukemia. 2016 Sep;30(9):1816-23. doi: 10.1038/leu.2016.164. Epub 2016 Jun 10. Leukemia. 2016. PMID: 27285583 Free PMC article. Review.
-
Transcriptome analysis of the binucleate ciliate Tetrahymena thermophila with asynchronous nuclear cell cycles.Mol Biol Cell. 2023 Feb 1;34(2):rs1. doi: 10.1091/mbc.E22-08-0326. Epub 2022 Dec 7. Mol Biol Cell. 2023. PMID: 36475712 Free PMC article.
References
-
- Altafaj X, Dierssen M, Baamonde C, Marti E, Visa J, Guimera J, Oset M, Gonzalez JR, Florez J, Fillat C, et al. 2001. Neurodevelopmental delay, motor abnormalities and cognitive deficits in transgenic mice overexpressing Dyrk1A (minibrain), a murine model of Down's syndrome. Hum Mol Genet 10: 1915–1923 - PubMed
-
- Aranda S, Laguna A, de la Luna S 2010. DYRK family of protein kinases: evolutionary relationships, biochemical properties, and functional roles. FASEB J 25: 449–462 - PubMed
-
- Beall EL, Manak JR, Zhou S, Bell M, Lipsick JS, Botchan MR 2002. Role for a Drosophila Myb-containing protein complex in site-specific DNA replication. Nature 420: 833–837 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials