

Phosphorylation-dependent regulation of the F-BAR protein Hof1 during cytokinesis
- PMID: 21498574
- PMCID: PMC3078711
- DOI: 10.1101/gad.622411
Phosphorylation-dependent regulation of the F-BAR protein Hof1 during cytokinesis
Abstract
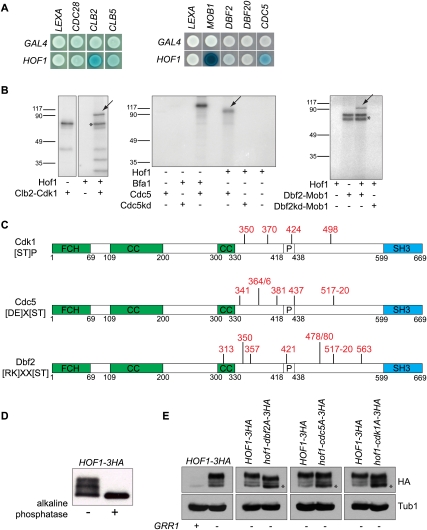
Spatial and timely coordination of cytokinesis is crucial for the maintenance of organelle inheritance and genome integrity. The mitotic exit network (MEN) pathway controls both the timely initiation of mitotic exit and cytokinesis in budding yeast. Here we identified the conserved F-BAR protein Hof1 as a substrate of the MEN kinase complex Dbf2-Mob1 during cytokinesis. We show that polo-like kinase Cdc5 first phosphorylates Hof1 to allow subsequent phosphorylation by Dbf2-Mob1. This releases Hof1 from the septin ring and facilitates Hof1 binding to the medial actomyosin ring (AMR), where Hof1 promotes AMR contraction and membrane ingression. Domain structure analysis established that the central, unstructured, region of Hof1, named the ring localization sequence (RLS), is sufficient to mediate Hof1's binding to the medial ring in a cell cycle-dependent manner. Genetic and functional data support a model in which Dbf2-Mob1 regulates Hof1 by inducing domain rearrangements, leading to the exposure of the Hof1 RLS domain during telophase.
Figures







Similar articles
-
Dual function of the NDR-kinase Dbf2 in the regulation of the F-BAR protein Hof1 during cytokinesis.Mol Biol Cell. 2013 May;24(9):1290-304. doi: 10.1091/mbc.E12-08-0608. Epub 2013 Feb 27. Mol Biol Cell. 2013. PMID: 23447700 Free PMC article.
-
Phosphorylation of the F-BAR protein Hof1 drives septin ring splitting in budding yeast.Nat Commun. 2024 Apr 22;15(1):3383. doi: 10.1038/s41467-024-47709-3. Nat Commun. 2024. PMID: 38649354 Free PMC article.
-
Mutation of Hof1 PEST motif phosphorylation sites leads to retention of Hof1 at the bud neck and a decrease in the rate of myosin contraction.Cell Biol Int. 2013 Apr;37(4):314-25. doi: 10.1002/cbin.10042. Epub 2013 Jan 29. Cell Biol Int. 2013. PMID: 23359466
-
Protein kinases in mitotic phosphorylation of budding yeast CENP-A.Curr Genet. 2019 Dec;65(6):1325-1332. doi: 10.1007/s00294-019-00997-5. Epub 2019 May 22. Curr Genet. 2019. PMID: 31119371 Review.
-
Central Role of the Actomyosin Ring in Coordinating Cytokinesis Steps in Budding Yeast.J Fungi (Basel). 2024 Sep 21;10(9):662. doi: 10.3390/jof10090662. J Fungi (Basel). 2024. PMID: 39330421 Free PMC article. Review.
Cited by
-
The Mitotic Exit Network and Cdc14 phosphatase initiate cytokinesis by counteracting CDK phosphorylations and blocking polarised growth.EMBO J. 2012 Aug 29;31(17):3620-34. doi: 10.1038/emboj.2012.224. Epub 2012 Aug 7. EMBO J. 2012. PMID: 22872148 Free PMC article.
-
Cell polarization and cytokinesis in budding yeast.Genetics. 2012 Jun;191(2):347-87. doi: 10.1534/genetics.111.132886. Genetics. 2012. PMID: 22701052 Free PMC article. Review.
-
The F-BAR protein Hof1 tunes formin activity to sculpt actin cables during polarized growth.Mol Biol Cell. 2014 Jun;25(11):1730-43. doi: 10.1091/mbc.E14-03-0850. Epub 2014 Apr 9. Mol Biol Cell. 2014. PMID: 24719456 Free PMC article.
-
Functions and regulation of the Polo-like kinase Cdc5 in the absence and presence of DNA damage.Curr Genet. 2018 Feb;64(1):87-96. doi: 10.1007/s00294-017-0727-2. Epub 2017 Aug 2. Curr Genet. 2018. PMID: 28770345 Free PMC article. Review.
-
A dynamical model of the spindle position checkpoint.Mol Syst Biol. 2012 May 8;8:582. doi: 10.1038/msb.2012.15. Mol Syst Biol. 2012. PMID: 22580890 Free PMC article.
References
-
- Aspenstrom P 2009. Roles of F-BAR/PCH proteins in the regulation of membrane dynamics and actin reorganization. Int Rev Cell Mol Biol 272: 1–31 - PubMed
-
- Aspenstrom P, Fransson A, Richnau N 2006. Pombe Cdc15 homology proteins: regulators of membrane dynamics and the actin cytoskeleton. Trends Biochem Sci 31: 670–679 - PubMed
-
- Balasubramanian MK, Bi E, Glotzer M 2004. Comparative analysis of cytokinesis in budding yeast, fission yeast and animal cells. Curr Biol 14: R806–R818 doi: 10.1016/j.cub.2004.09.022 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases