

Mobile gibberellin directly stimulates Arabidopsis hypocotyl xylem expansion
- PMID: 21498678
- PMCID: PMC3101547
- DOI: 10.1105/tpc.111.084020
Mobile gibberellin directly stimulates Arabidopsis hypocotyl xylem expansion
Abstract
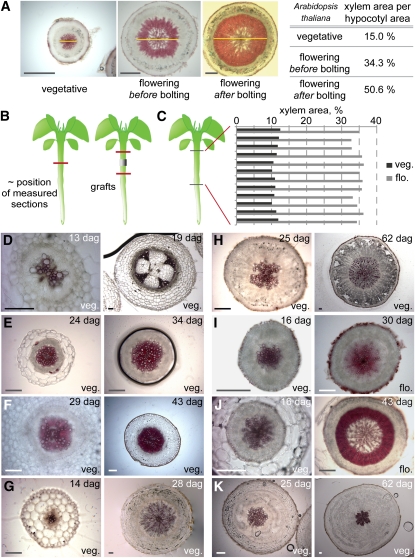
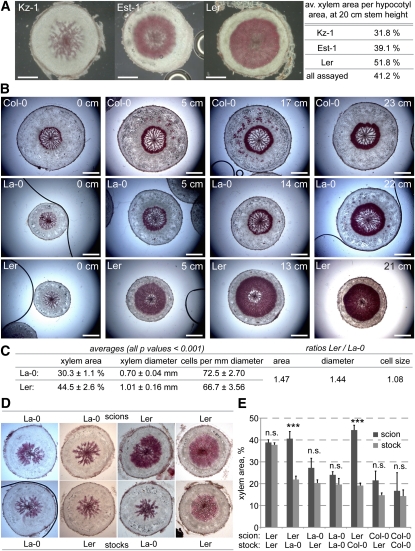
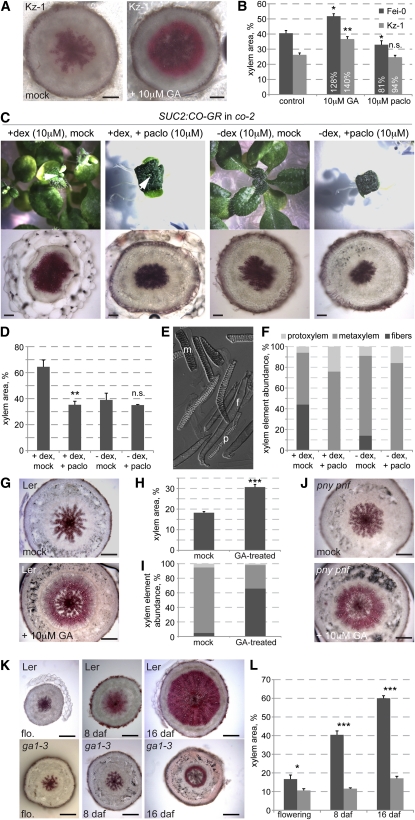
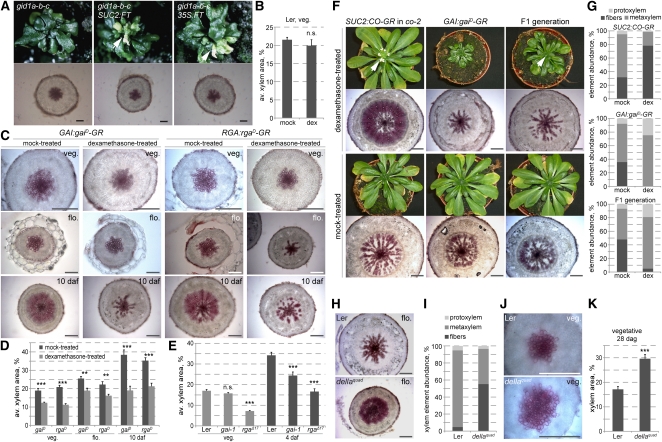
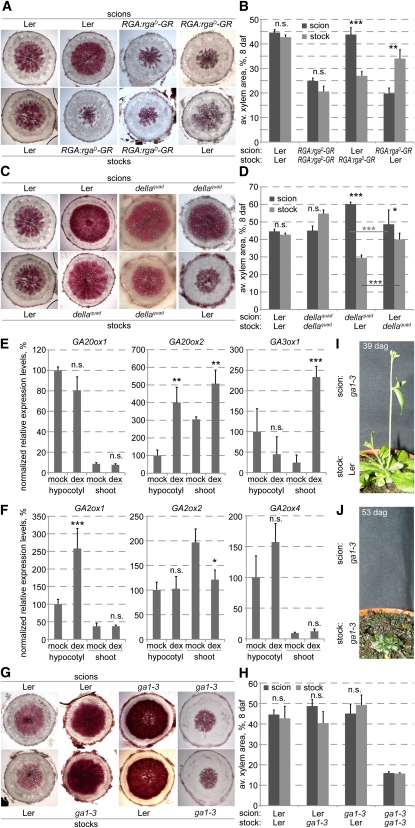
Secondary growth of the vasculature results in the thickening of plant structures and continuously produces xylem tissue, the major biological carbon sink. Little is known about the developmental control of this quantitative trait, which displays two distinct phases in Arabidopsis thaliana hypocotyls. The later phase of accelerated xylem expansion resembles the secondary growth of trees and is triggered upon flowering by an unknown, shoot-derived signal. We found that flowering-dependent hypocotyl xylem expansion is a general feature of herbaceous plants with a rosette growth habit. Flowering induction is sufficient to trigger xylem expansion in Arabidopsis. By contrast, neither flower formation nor elongation of the main inflorescence is required. Xylem expansion also does not depend on any particular flowering time pathway or absolute age. Through analyses of natural genetic variation, we found that ERECTA acts locally to restrict xylem expansion downstream of the gibberellin (GA) pathway. Investigations of mutant and transgenic plants indicate that GA and its signaling pathway are both necessary and sufficient to directly trigger enhanced xylogenesis. Impaired GA signaling did not affect xylem expansion systemically, suggesting that it acts downstream of the mobile cue. By contrast, the GA effect was graft transmissible, suggesting that GA itself is the mobile shoot-derived signal.
Figures
References
-
- Achard P., Cheng H., De Grauwe L., Decat J., Schoutteten H., Moritz T., Van Der Straeten D., Peng J., Harberd N.P. (2006). Integration of plant responses to environmentally activated phytohormonal signals. Science 311: 91–94 - PubMed
-
- Berleth T., Mattsson J., Hardtke C.S. (2000). Vascular continuity and auxin signals. Trends Plant Sci. 5: 387–393 - PubMed
-
- Björklund S., Antti H., Uddestrand I., Moritz T., Sundberg B. (2007). Cross-talk between gibberellin and auxin in development of Populus wood: Gibberellin stimulates polar auxin transport and has a common transcriptome with auxin. Plant J. 52: 499–511 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
