

A Drosophila model for alcohol reward
- PMID: 21499254
- PMCID: PMC4249630
- DOI: 10.1038/nn.2805
A Drosophila model for alcohol reward
Abstract
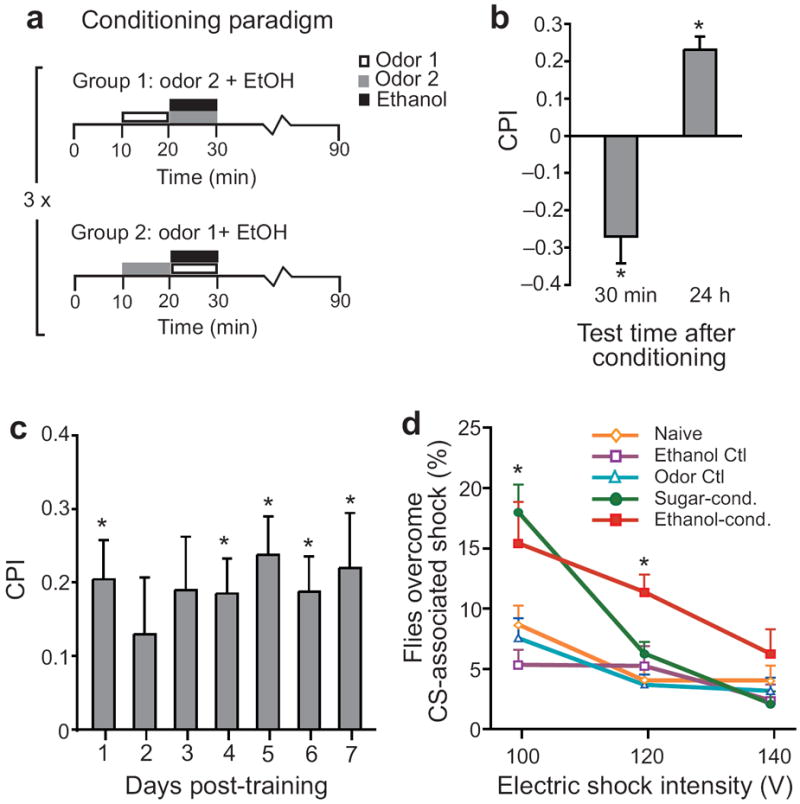
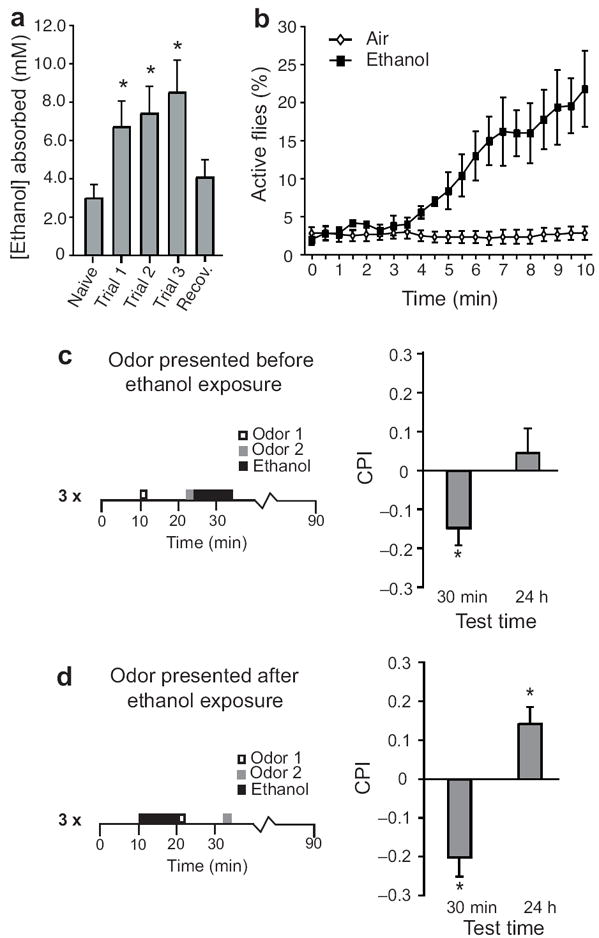
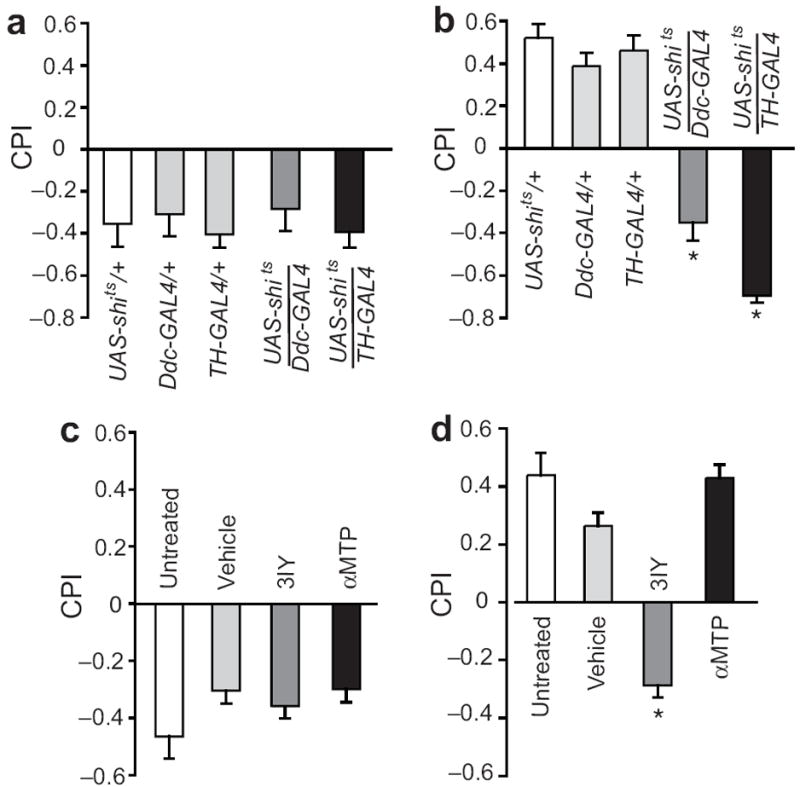
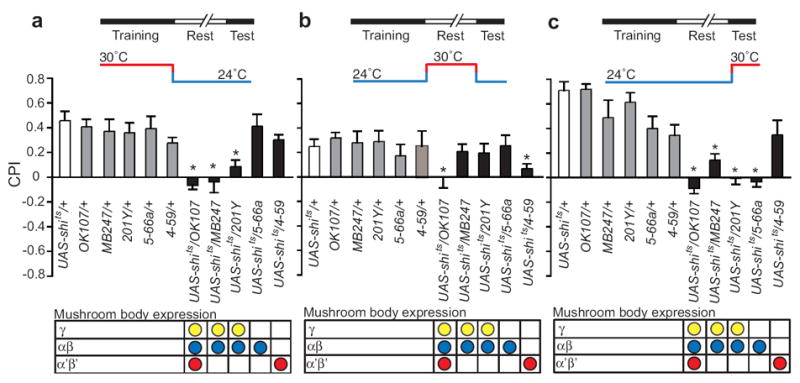
The rewarding properties of drugs contribute to the development of abuse and addiction. We developed a new assay for investigating the motivational properties of ethanol in the genetically tractable model Drosophila melanogaster. Flies learned to associate cues with ethanol intoxication and, although transiently aversive, the experience led to a long-lasting attraction for the ethanol-paired cue, implying that intoxication is rewarding. Temporally blocking transmission in dopaminergic neurons revealed that flies require activation of these neurons to express, but not develop, conditioned preference for ethanol-associated cues. Moreover, flies acquired, consolidated and retrieved these rewarding memories using distinct sets of neurons in the mushroom body. Finally, mutations in scabrous, encoding a fibrinogen-related peptide that regulates Notch signaling, disrupted the formation of memories for ethanol reward. Our results thus establish that Drosophila can be useful for understanding the molecular, genetic and neural mechanisms underling the rewarding properties of ethanol.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
-
- Hyman SE, Malenka RC, Nestler EJ. Neural mechanisms of addiction: The role of reward-related learning and memory. Annu Rev Neurosci. 2006;29:565–598. - PubMed
-
- Tzschentke TM. Measuring reward with the conditioned place preference (CPP) paradigm: update of the last decade. Addict Biol. 2007;12:227–262. - PubMed
-
- Baker NE, Mlodzik M, Rubin GM. Spacing differentiation in the developing Drosophila eye: A fibrinogen-related lateral inhibitor encoded by scabrous. Science. 1990;250:1370–1377. - PubMed
-
- Powell PA, Wesley C, Spencer S, Cagan RL. Scabrous complexes with Notch to mediate boundary formation. Nature. 2001;409:626–630. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
