

Control of nuclear receptor function by local chromatin structure
- PMID: 21501385
- PMCID: PMC6319360
- DOI: 10.1111/j.1742-4658.2011.08126.x
Control of nuclear receptor function by local chromatin structure
Abstract
Steroid hormone receptors regulate gene transcription in a highly tissue-specific manner. The local chromatin structure underlying promoters and hormone response elements is a major component involved in controlling these highly restricted expression patterns. Chromatin remodeling complexes, as well as histone and DNA modifying enzymes, are directed to gene-specific regions and create permissive or repressive chromatin environments. These structures further enable proper communication between transcription factors, co-regulators and basic transcription machinery. The regulatory elements active at target genes can be either constitutively accessible to receptors or subject to rapid receptor-dependent modification. The chromatin states responsible for these processes are in turn determined during development and differentiation. Thus access of regulatory factors to elements in chromatin provides a major level of cell selective regulation.
Journal compilation © 2011 FEBS. No claim to original US government works.
Figures
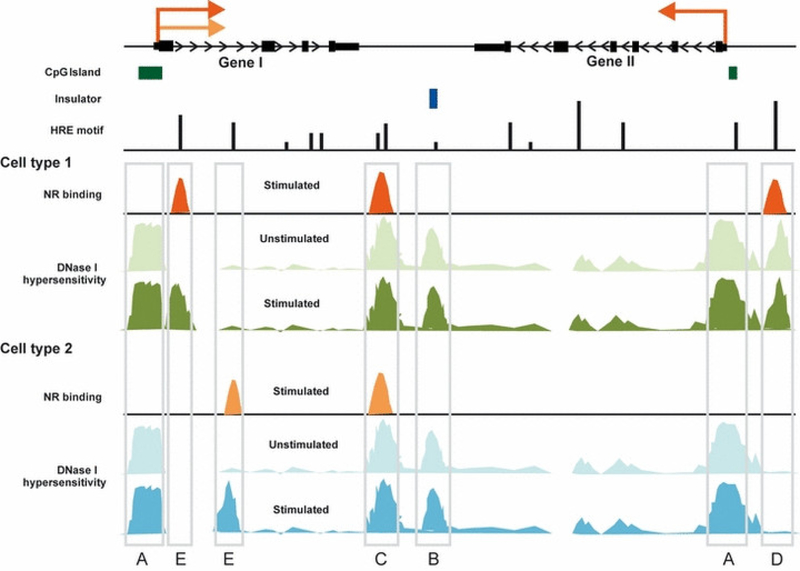
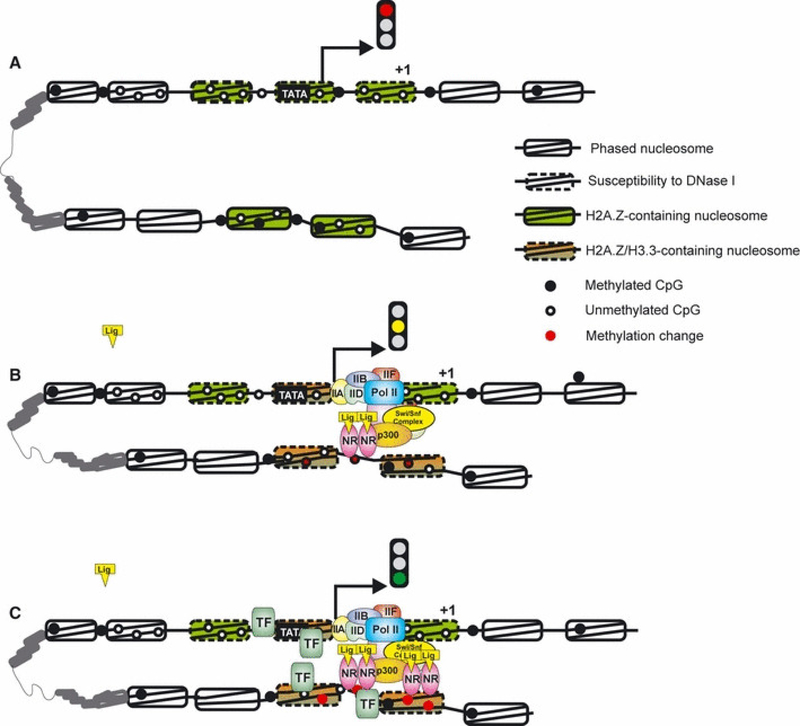
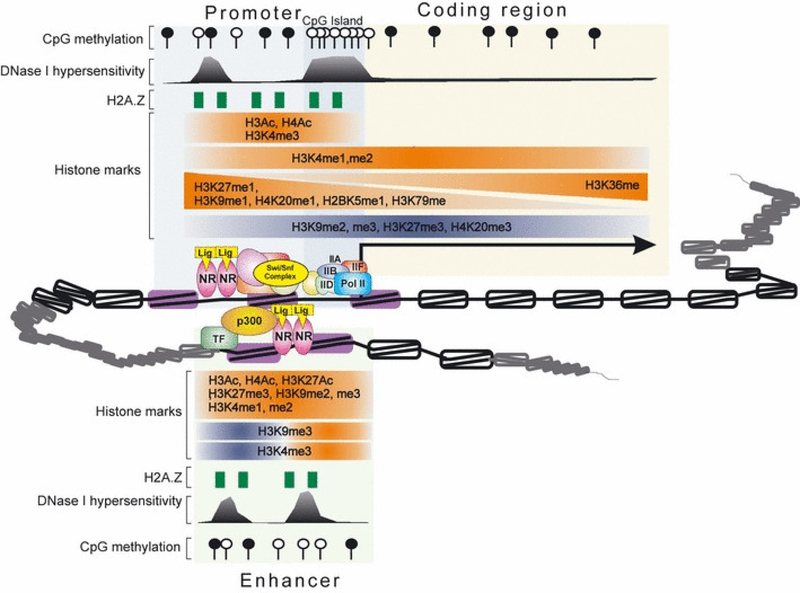
References
-
- Beato M & Klug J (2000) Steroid hormone receptors: an update. Hum Reprod Update 6, 225–236. - PubMed
-
- Cockerill PN (2011) Structure and function of active chromatin and DNase I hypersensitive sites. FEBS J 278, 2182–2210. - PubMed
-
- Bednar J & Dimitrov S (2011) Chromatin under mechanical stress: from single 30 nm fibers to single nucleosomes. FEBS J 278, 2231–2243. - PubMed
-
- Richmond TJ, Finch JT, Rushton B, Rhodes D & Klug A (1984) Structure of the nucleosome core particle at 7Å resolution. Nature 311, 532–537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
