

Assembly and functions of heterochromatin in the fission yeast genome
- PMID: 21502415
- PMCID: PMC6309827
- DOI: 10.1101/sqb.2010.75.055
Assembly and functions of heterochromatin in the fission yeast genome
Abstract
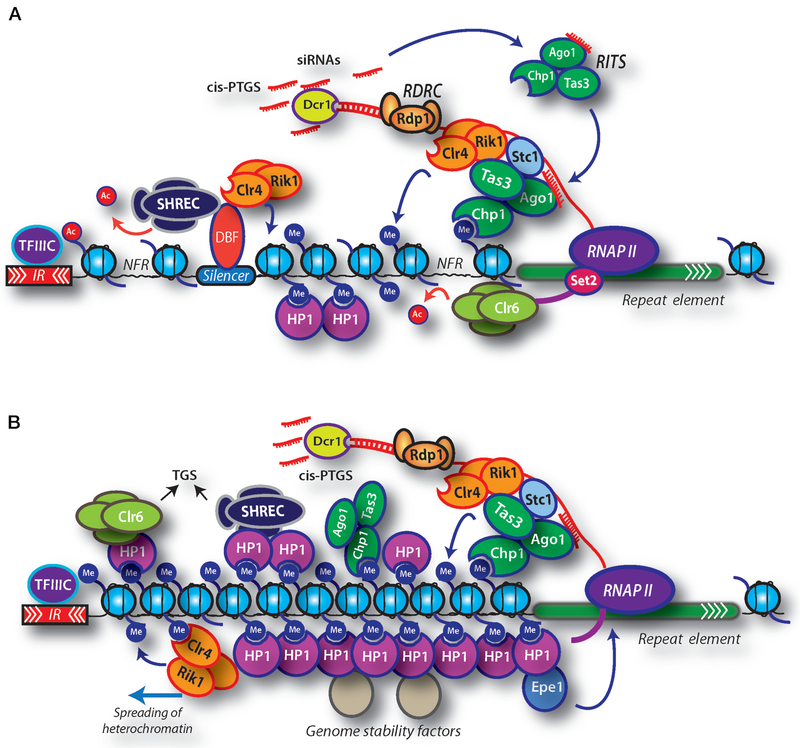
In eukaryotic genomes, heterochromatin regulates various chromosomal processes including suppression of transcription and illegitimate recombination as well as proper segregation of chromosomes during cell division. Recent studies using the fission yeast Schizosaccharomyces pombe model system have revealed a complex interplay among RNA polymerase II transcription, RNAi machinery, and factors involved in posttranslational modifications of histones that are critical for the assembly and maintenance of heterochromatin. Heterochromatin proteins targeted to specific sites in the genome can spread across extended chromosomal domains and mediate epigenetic genome control by providing a recruitment platform for various factors including chromatin-modifying activities. In this chapter, we discuss mechanisms of heterochromatin assembly in fission yeast and highlight emerging evidence suggesting the involvement of heterochromatin factors in the suppression of noncoding RNAs across the genome.
Figures
Similar articles
-
RNAi-mediated chromatin silencing in fission yeast.Curr Top Microbiol Immunol. 2008;320:157-83. doi: 10.1007/978-3-540-75157-1_8. Curr Top Microbiol Immunol. 2008. PMID: 18268844 Review.
-
Unique roles for histone H3K9me states in RNAi and heritable silencing of transcription.Nature. 2017 Jul 27;547(7664):463-467. doi: 10.1038/nature23267. Epub 2017 Jun 22. Nature. 2017. PMID: 28682306 Free PMC article.
-
Epigenetic Regulation of Chromatin States in Schizosaccharomyces pombe.Cold Spring Harb Perspect Biol. 2015 Jul 1;7(7):a018770. doi: 10.1101/cshperspect.a018770. Cold Spring Harb Perspect Biol. 2015. PMID: 26134317 Free PMC article. Review.
-
RNAi-mediated heterochromatin assembly in fission yeast.Cold Spring Harb Symp Quant Biol. 2006;71:487-96. doi: 10.1101/sqb.2006.71.059. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381331 Review.
-
Different means, same end-heterochromatin formation by RNAi and RNAi-independent RNA processing factors in fission yeast.Curr Opin Genet Dev. 2012 Apr;22(2):156-63. doi: 10.1016/j.gde.2011.12.004. Epub 2012 Jan 11. Curr Opin Genet Dev. 2012. PMID: 22243696 Free PMC article. Review.
Cited by
-
Chromosome segregation and organization are targets of 5'-Fluorouracil in eukaryotic cells.Cell Cycle. 2015;14(2):206-18. doi: 10.4161/15384101.2014.974425. Cell Cycle. 2015. PMID: 25483073 Free PMC article.
-
Endogenous nuclear RNAi mediates behavioral adaptation to odor.Cell. 2013 Aug 29;154(5):1010-1022. doi: 10.1016/j.cell.2013.08.006. Cell. 2013. PMID: 23993094 Free PMC article.
-
A Novel Epigenetic Silencing Pathway Involving the Highly Conserved 5'-3' Exoribonuclease Dhp1/Rat1/Xrn2 in Schizosaccharomyces pombe.PLoS Genet. 2016 Feb 18;12(2):e1005873. doi: 10.1371/journal.pgen.1005873. eCollection 2016 Feb. PLoS Genet. 2016. PMID: 26889830 Free PMC article.
-
Biochemical Basis for Distinct Roles of the Heterochromatin Proteins Swi6 and Chp2.J Mol Biol. 2017 Nov 24;429(23):3666-3677. doi: 10.1016/j.jmb.2017.09.012. Epub 2017 Sep 20. J Mol Biol. 2017. PMID: 28942089 Free PMC article.
-
Enforcing silencing: dynamic HP1 complexes in Neurospora.Nat Struct Mol Biol. 2012 May 3;19(5):465-7. doi: 10.1038/nsmb.2291. Nat Struct Mol Biol. 2012. PMID: 22551706
References
-
- Allshire RC. 1995. Elements of chromosome structure and function in fission yeast. Semin Cell Biol 6: 55–64. - PubMed
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T. 2001. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 410: 120–124. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources