

New investigations around CYP11A1 and its possible involvement in an androstenone QTL characterised in Large White pigs
- PMID: 21504607
- PMCID: PMC3098772
- DOI: 10.1186/1297-9686-43-15
New investigations around CYP11A1 and its possible involvement in an androstenone QTL characterised in Large White pigs
Abstract
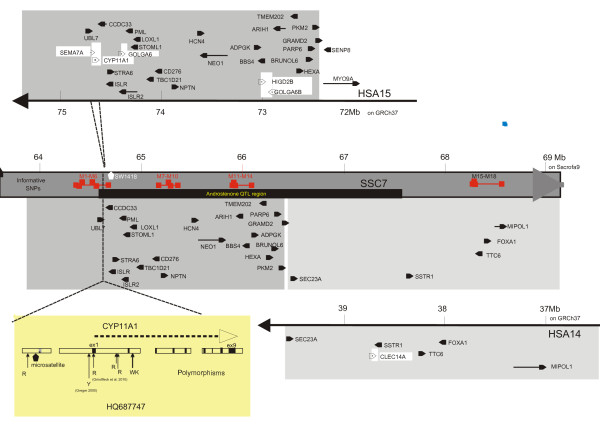
Background: Previously, in boars with extreme androstenone levels, differential expression of the CYP11A1 gene in the testes has been characterised. CYP11A1 is located in a region where a QTL influencing boar fat androstenone levels has been detected in a Large White pig population. Clarifying the role of CYP11A1 in boar taint is important because it catalyses the initial step of androstenone synthesis and also of steroid synthesis.
Results: A genome-wide association study located CYP11A1 at approximately 1300 kb upstream from SNP H3GA0021967, defining the centre of the region containing the QTL for androstenone variation. In this study, we partially sequenced the CYP11A1 gene and identified several new single nucleotide polymorphisms (SNP) within it. Characterisation of one animal, heterozygous for CYP11A1 testicular expression but homozygous for a haplotype of a large region containing CYP11A1, revealed that variation of CYP11A1 expression is probably regulated by a mutation located downstream from the SNP H3GA0021967. We analysed CYP11A1 expression in LW families according to haplotypes of the QTL region's centre. Effects of haplotypes on CYP11A1 expression and on androstenone accumulation were not concordant.
Conclusion: This study shows that testicular expression of CYP11A1 is not solely responsible for the QTL influencing boar fat androstenone levels. As a conclusion, we propose to refute the hypothesis that a single mutation located near the centre of the QTL region could control androstenone accumulation in fat by regulating the CYP11A1 expression.
Figures
Similar articles
-
On the relationship between an Asian haplotype on chromosome 6 that reduces androstenone levels in boars and the differential expression of SULT2A1 in the testis.BMC Genet. 2014 Jan 9;15:4. doi: 10.1186/1471-2156-15-4. BMC Genet. 2014. PMID: 24405739 Free PMC article.
-
Genome-wide association scan and phased haplotype construction for quantitative trait loci affecting boar taint in three pig breeds.BMC Genomics. 2012 Jan 13;13:22. doi: 10.1186/1471-2164-13-22. BMC Genomics. 2012. PMID: 22244367 Free PMC article.
-
Boar taint in entire male pigs: a genomewide association study for direct and indirect genetic effects on androstenone.J Anim Sci. 2014 Oct;92(10):4319-28. doi: 10.2527/jas.2014-7863. Epub 2014 Aug 22. J Anim Sci. 2014. PMID: 25149343
-
Genetic and metabolic aspects of androstenone and skatole deposition in pig adipose tissue: a review.Genet Sel Evol. 2008 Jan-Feb;40(1):129-43. doi: 10.1186/1297-9686-40-1-129. Epub 2007 Dec 21. Genet Sel Evol. 2008. PMID: 18096119 Free PMC article. Review.
-
Recent genetic advances on boar taint reduction as an alternative to castration: a review.J Appl Genet. 2021 Feb;62(1):137-150. doi: 10.1007/s13353-020-00598-w. Epub 2021 Jan 6. J Appl Genet. 2021. PMID: 33405214 Free PMC article. Review.
Cited by
-
Two new structural mutations in the 5' region of the ASIP gene cause diluted feather color phenotypes in Japanese quail.Genet Sel Evol. 2019 Apr 15;51(1):12. doi: 10.1186/s12711-019-0458-6. Genet Sel Evol. 2019. PMID: 30987584 Free PMC article.
-
Analysis of the genetics of boar taint reveals both single SNPs and regional effects.BMC Genomics. 2014 Jun 3;15(1):424. doi: 10.1186/1471-2164-15-424. BMC Genomics. 2014. PMID: 24894739 Free PMC article.
-
Efficiency of genomic prediction for boar taint reduction in Danish Landrace pigs.Anim Genet. 2015 Dec;46(6):607-16. doi: 10.1111/age.12369. Epub 2015 Oct 9. Anim Genet. 2015. PMID: 26449733 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
