

Zinc effects on NMDA receptor gating kinetics
- PMID: 21504727
- PMCID: PMC3077707
- DOI: 10.1016/j.bpj.2011.02.042
Zinc effects on NMDA receptor gating kinetics
Abstract
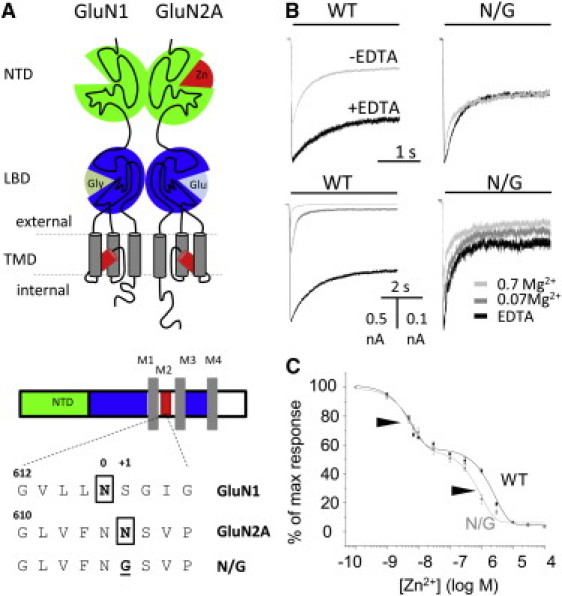
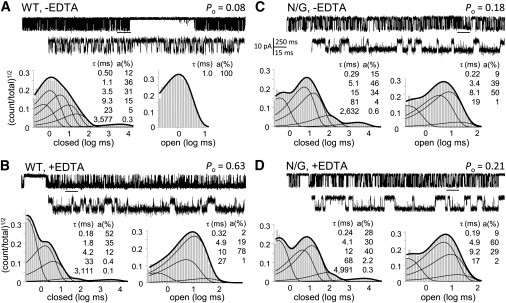
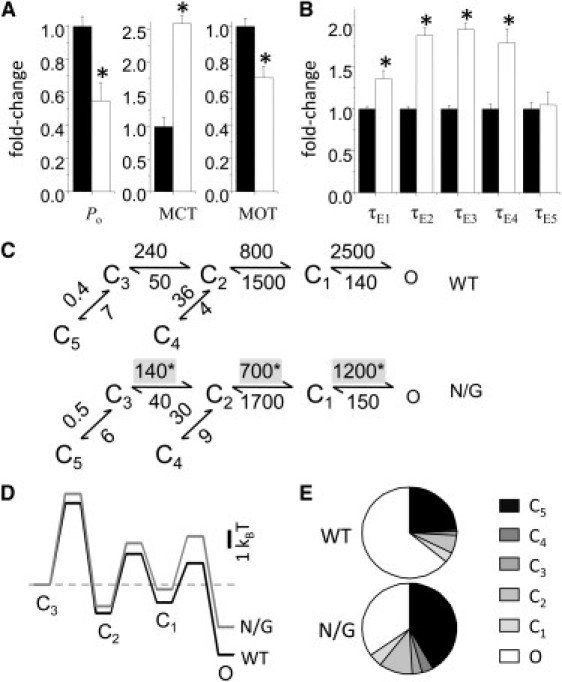
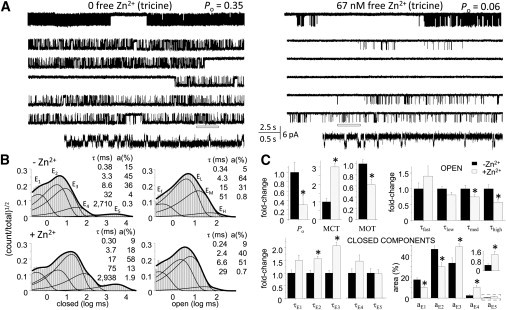
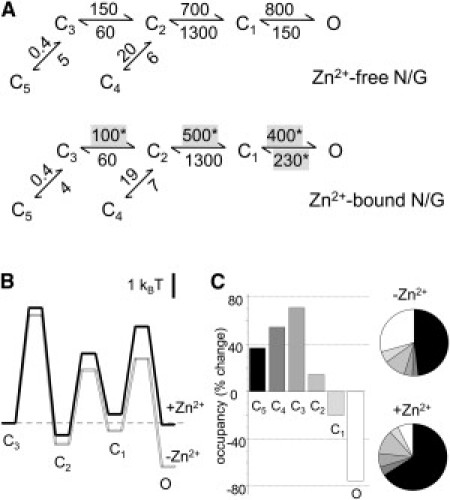
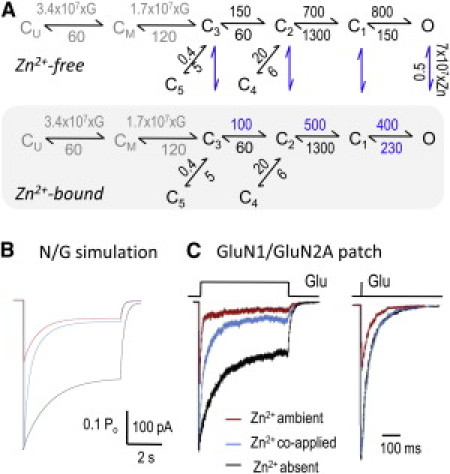
Zinc accumulates in the synaptic vesicles of certain glutamatergic forebrain neurons and modulates neuronal excitability and synaptic plasticity by multiple poorly understood mechanisms. Zinc directly inhibits NMDA-sensitive glutamate-gated channels by two separate mechanisms: high-affinity binding to N-terminal domains of GluN2A subunits reduces channel open probability, and low-affinity voltage-dependent binding to pore-lining residues blocks the channel. Insight into the high-affinity allosteric effect has been hampered by the receptor's complex gating; multiple, sometimes coupled, modulatory mechanisms; and practical difficulties in avoiding transient block by residual Mg(2+). To sidestep these challenges, we examined how nanomolar zinc concentrations changed the gating kinetics of individual block-resistant receptors. We found that block-insensitive channels had lower intrinsic open probabilities but retained high sensitivity to zinc inhibition. Binding of zinc to these receptors resulted in longer closures and shorter openings within bursts of activity but had no effect on interburst intervals. Based on kinetic modeling of these data, we conclude that zinc-bound receptors have higher energy barriers to opening and less stable open states. We tested this model for its ability to predict zinc-dependent changes in macroscopic responses and to infer the impact of nanomolar zinc concentrations on synaptic currents mediated by 2A-type NMDA receptors.
Copyright © 2011 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Takeda A. Zinc homeostasis and functions of zinc in the brain. Biometals. 2001;14:343–351. - PubMed
-
- Sensi S.L., Paoletti P., Sekler I. Zinc in the physiology and pathology of the CNS. Nat. Rev. Neurosci. 2009;10:780–791. - PubMed
-
- Frederickson C.J., Bush A.I. Synaptically released zinc: physiological functions and pathological effects. Biometals. 2001;14:353–366. - PubMed
-
- Smart T.G., Hosie A.M., Miller P.S. Zn2+ ions: modulators of excitatory and inhibitory synaptic activity. Neuroscientist. 2004;10:432–442. - PubMed
-
- Assaf S.Y., Chung S.H. Release of endogenous Zn2+ from brain tissue during activity. Nature. 1984;308:734–736. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
