

Convergent evolution in biosynthesis of cyanogenic defence compounds in plants and insects
- PMID: 21505429
- PMCID: PMC4354137
- DOI: 10.1038/ncomms1271
Convergent evolution in biosynthesis of cyanogenic defence compounds in plants and insects
Erratum in
- Nat Commun. 2012;3. doi:10.1038/ncomms1865
Abstract
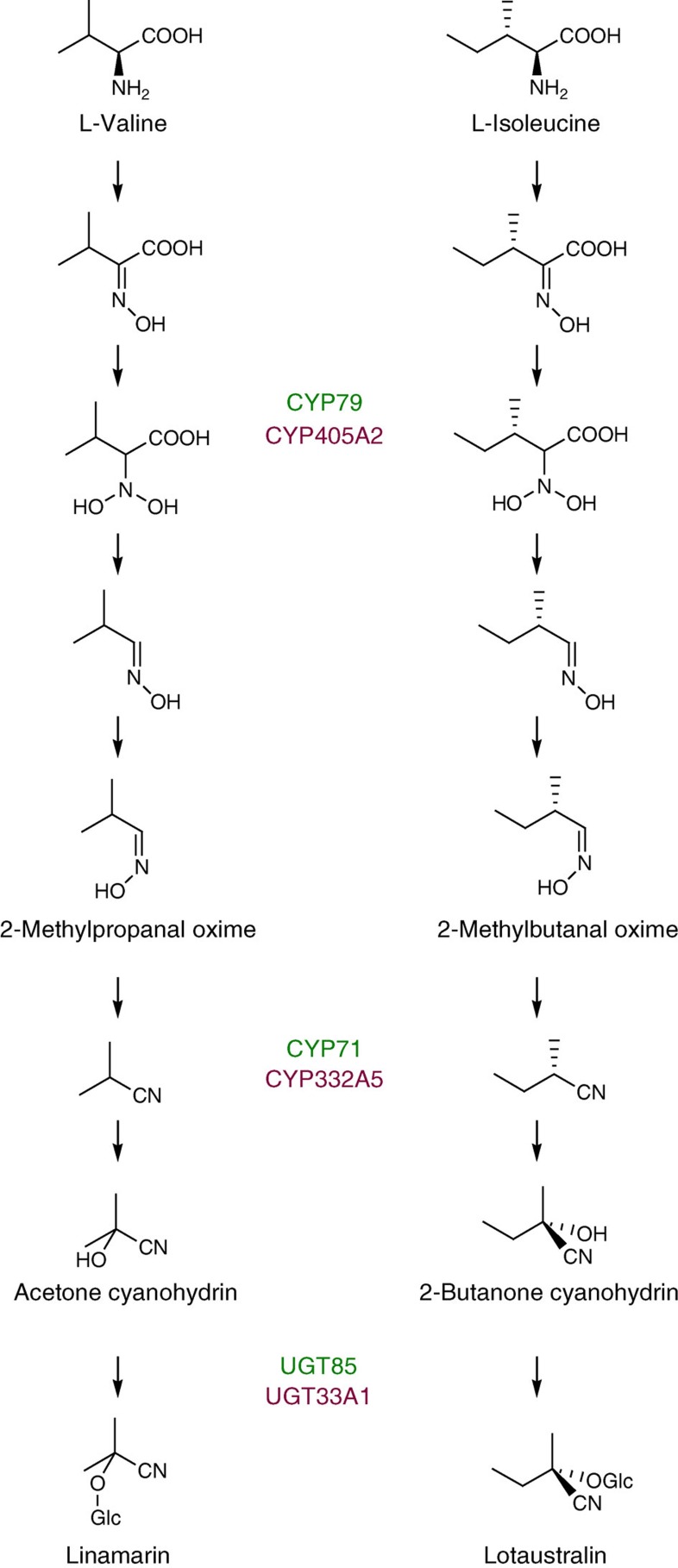
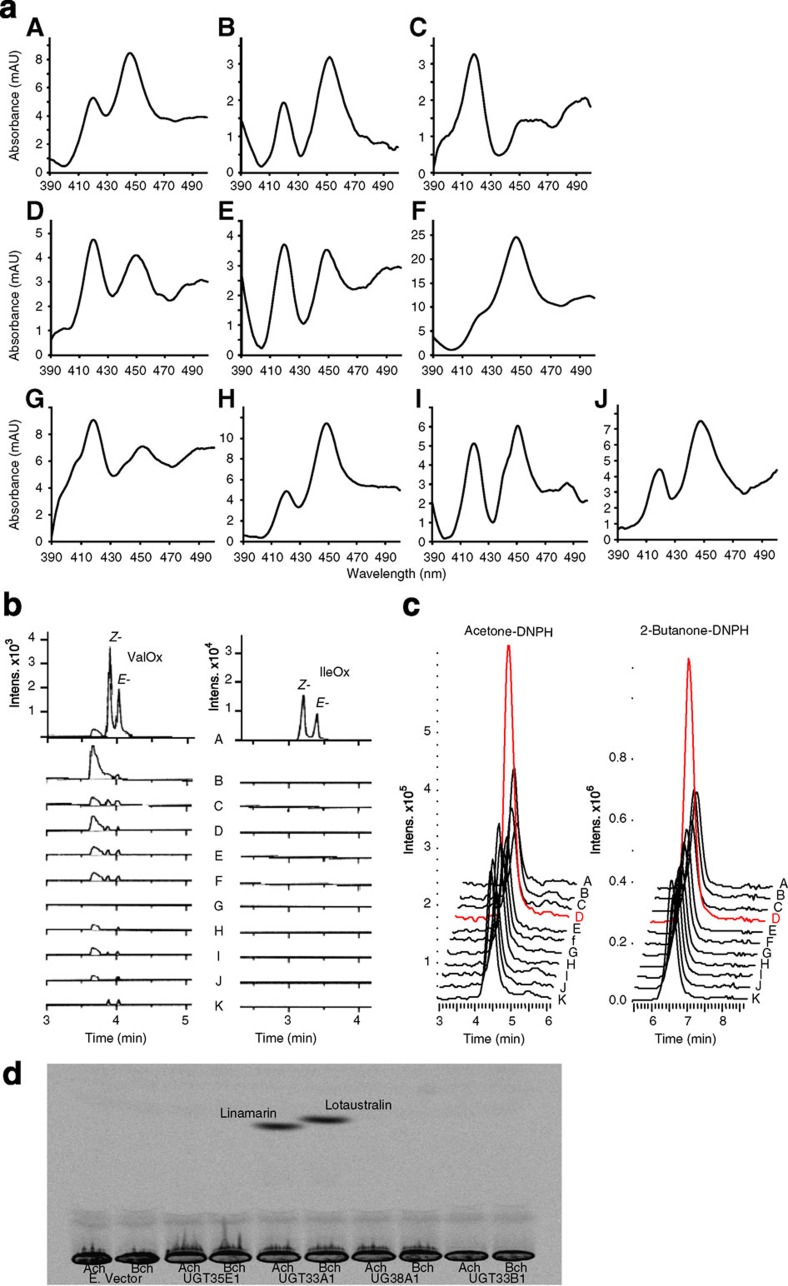
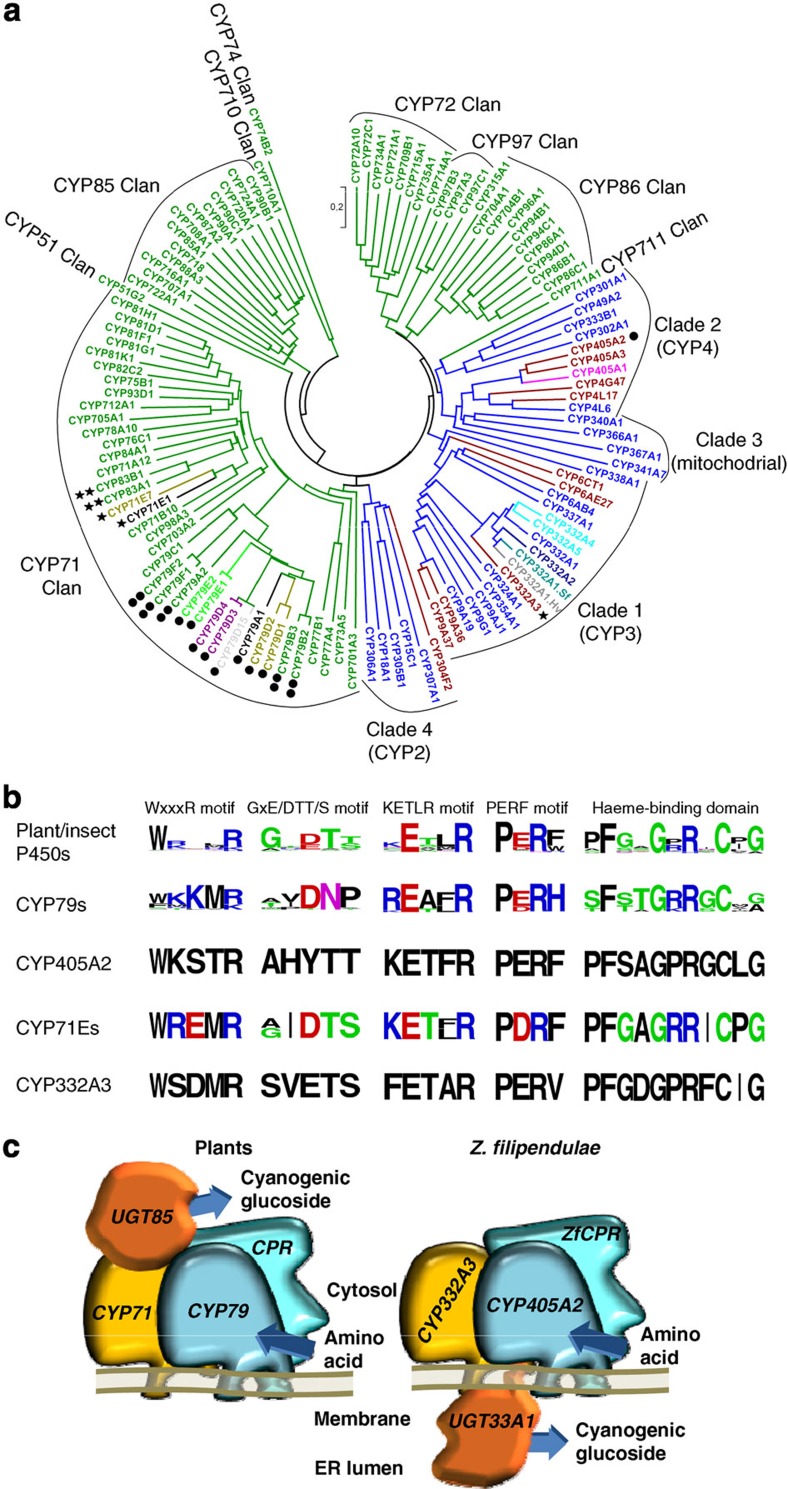
For more than 420 million years, plants, insects and their predators have co-evolved based on a chemical arms race including deployment of refined chemical defence systems by each player. Cyanogenic glucosides are produced by numerous plants and by some specialized insects and serve an important role as defence compounds in these intimate interactions. Burnet moth larvae are able to sequester cyanogenic glucosides from their food plant as well as to carry out de novo biosynthesis. Here we show that three genes (CYP405A2, CYP332A3 and UGT33A1) encode the entire biosynthetic pathway of cyanogenic glucosides in the Burnet moth Zygaena filipendulae. In both plants and insects, convergent evolution has led to two multifunctional P450 enzymes each catalysing unusual reactions and a glucosyl-transferase acting in sequence to catalyse cyanogenic glucoside formation. Thus, plants and insects have independently found a way to package a cyanide time bomb to fend off herbivores and predators.
Figures

Similar articles
-
454 pyrosequencing based transcriptome analysis of Zygaena filipendulae with focus on genes involved in biosynthesis of cyanogenic glucosides.BMC Genomics. 2009 Dec 2;10:574. doi: 10.1186/1471-2164-10-574. BMC Genomics. 2009. PMID: 19954531 Free PMC article.
-
Evolution of the Biosynthetic Pathway for Cyanogenic Glucosides in Lepidoptera.J Mol Evol. 2018 Jul;86(6):379-394. doi: 10.1007/s00239-018-9854-8. Epub 2018 Jul 4. J Mol Evol. 2018. PMID: 29974176
-
Cyanogenic glucosides in the biological warfare between plants and insects: the Burnet moth-Birdsfoot trefoil model system.Phytochemistry. 2011 Sep;72(13):1585-92. doi: 10.1016/j.phytochem.2011.02.023. Epub 2011 Mar 21. Phytochemistry. 2011. PMID: 21429539 Review.
-
Cyanogenesis in Arthropods: From Chemical Warfare to Nuptial Gifts.Insects. 2018 May 3;9(2):51. doi: 10.3390/insects9020051. Insects. 2018. PMID: 29751568 Free PMC article. Review.
-
Cyanogenesis in plants and arthropods.Phytochemistry. 2008 May;69(7):1457-68. doi: 10.1016/j.phytochem.2008.02.019. Epub 2008 Mar 18. Phytochemistry. 2008. PMID: 18353406 Review.
Cited by
-
Differential phenotypic and genetic expression of defence compounds in a plant-herbivore interaction along elevation.R Soc Open Sci. 2016 Sep 28;3(9):160226. doi: 10.1098/rsos.160226. eCollection 2016 Sep. R Soc Open Sci. 2016. PMID: 27703688 Free PMC article.
-
Phosphate solubilizing microorganisms isolated from medicinal plants improve growth of mint.PeerJ. 2022 Aug 17;10:e13782. doi: 10.7717/peerj.13782. eCollection 2022. PeerJ. 2022. PMID: 35996668 Free PMC article.
-
Pyrethroid activity-based probes for profiling cytochrome P450 activities associated with insecticide interactions.Proc Natl Acad Sci U S A. 2013 Dec 3;110(49):19766-71. doi: 10.1073/pnas.1320185110. Epub 2013 Nov 18. Proc Natl Acad Sci U S A. 2013. PMID: 24248381 Free PMC article.
-
New paths of cyanogenesis from enzymatic-promoted cleavage of β-cyanoglucosides are suggested by a mixed DFT/QTAIM approach.J Mol Model. 2019 Sep 3;25(9):295. doi: 10.1007/s00894-019-4170-9. J Mol Model. 2019. PMID: 31478108
-
Elucidation of the Amygdalin Pathway Reveals the Metabolic Basis of Bitter and Sweet Almonds (Prunus dulcis).Plant Physiol. 2018 Nov;178(3):1096-1111. doi: 10.1104/pp.18.00922. Epub 2018 Oct 8. Plant Physiol. 2018. PMID: 30297455 Free PMC article.
References
-
- Bak S., Kahn R. A., Nielsen H. L., Møller B. L. & Halkier B. A. Cloning of three A type cytochromes P450, CYP71E1, CYP98, and CYP99 from Sorghum bicolor (L.) Moench by a PCR approach and identification by expression in Escherichia coli of CYP71E1 as a multifunctional cytochrome P450 in the biosynthesis of the cyanogenic glucoside dhurrin. Plant Mol. Biol. 36, 393–405 (1998). - PubMed
-
- Halkier B. A., Nielsen H. L., Koch B. & Moller B. L. Purification and characterization of recombinant cytochrome P450TYR expressed at high levels in Escherichia coli. Arch. Biochem. Biophys. 322, 369–377 (1995). - PubMed
-
- Tattersall D. B. et al.. Resistance to a herbivore through engineered cyanogenic glucoside synthesis. Science 293, 1826–1828 (2001). - PubMed
-
- Jones P. R., Møller B. L. & Høj P. B. The UDP-glucose: p-hydroxymandelonitrile-O-glucosyltransferase that catalyzes the last step in synthesis of the cyanogenic glucoside dhurrin in Sorghum bicolor - isolation, cloning, heterologous expression, and substrate specificity. J. Biol. Chem. 274, 35483–35491 (1999). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
