

Rapid cell-surface prion protein conversion revealed using a novel cell system
- PMID: 21505437
- PMCID: PMC3104518
- DOI: 10.1038/ncomms1282
Rapid cell-surface prion protein conversion revealed using a novel cell system
Abstract
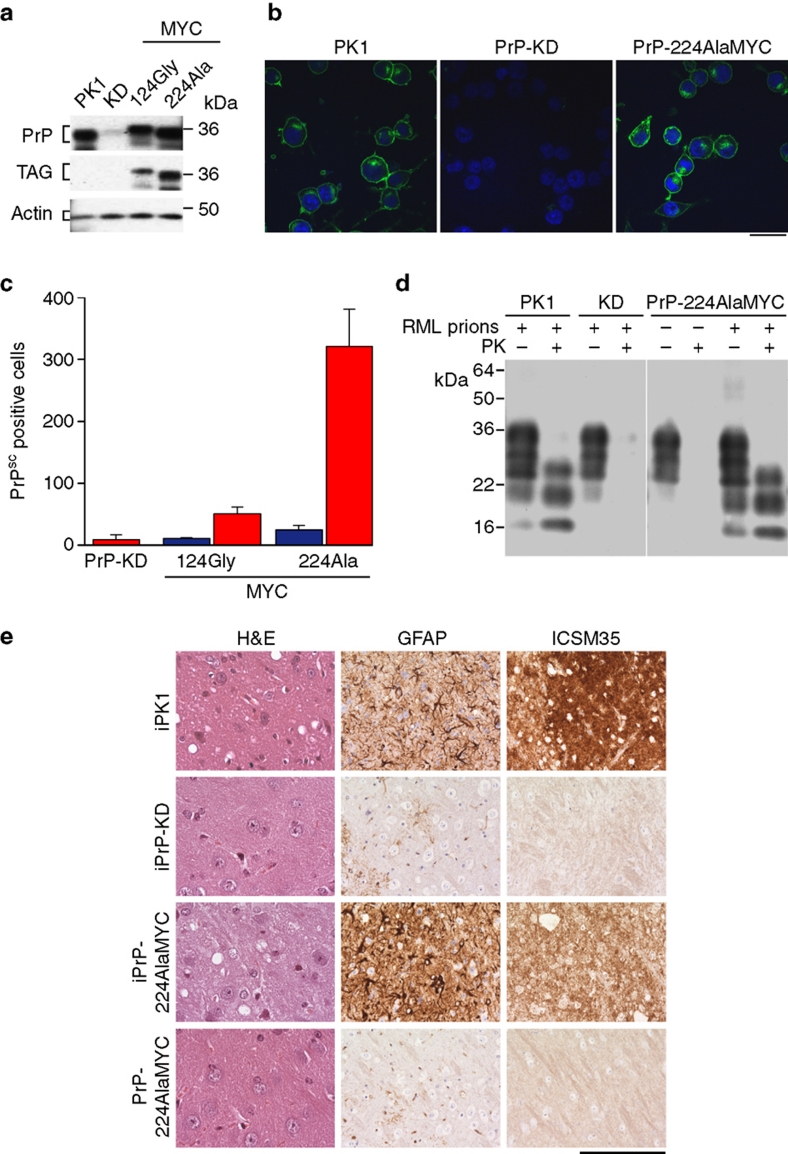
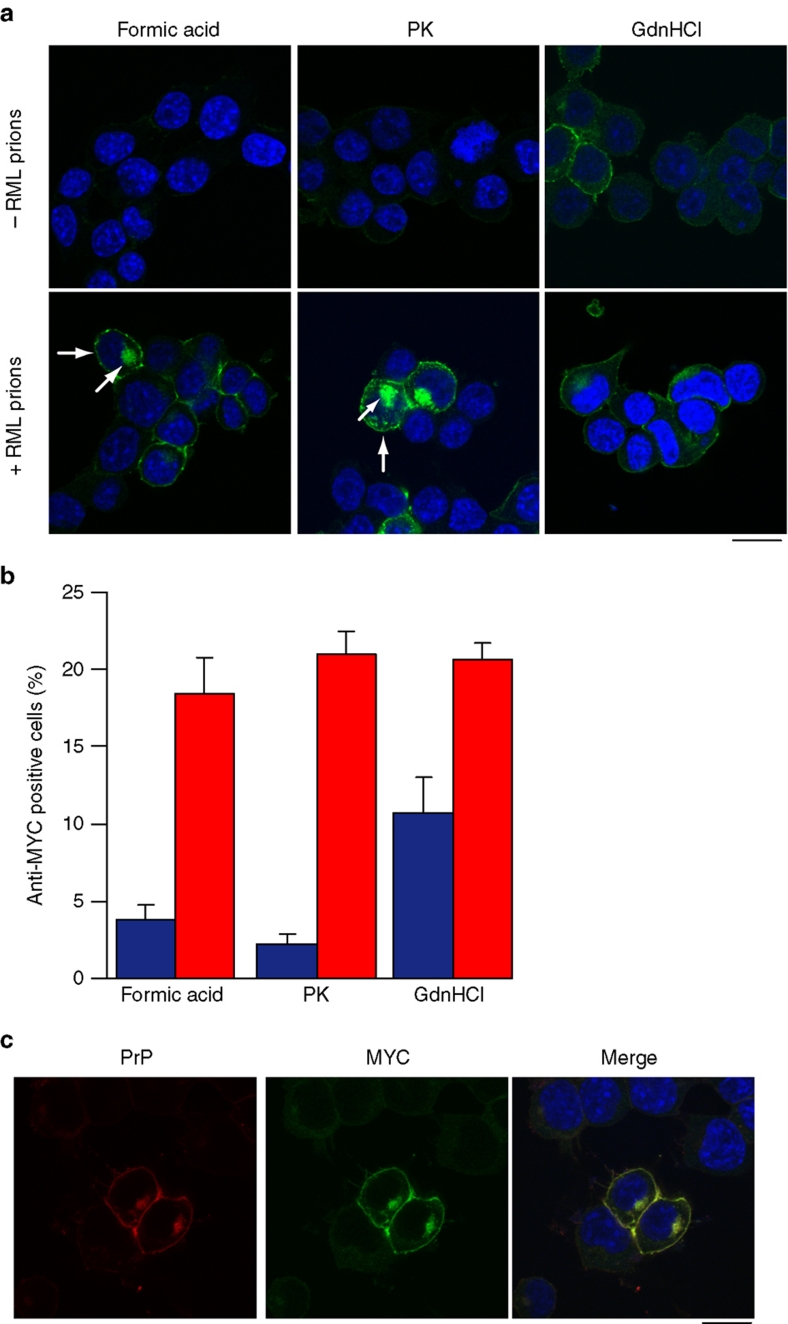
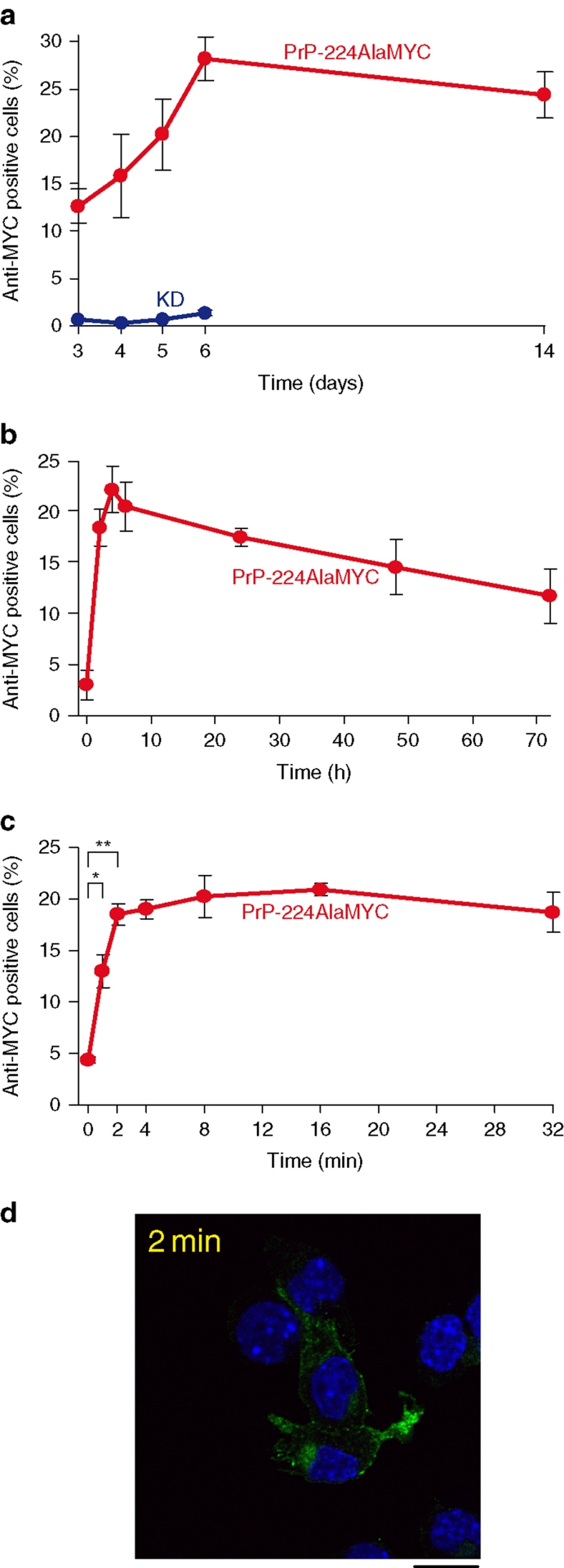
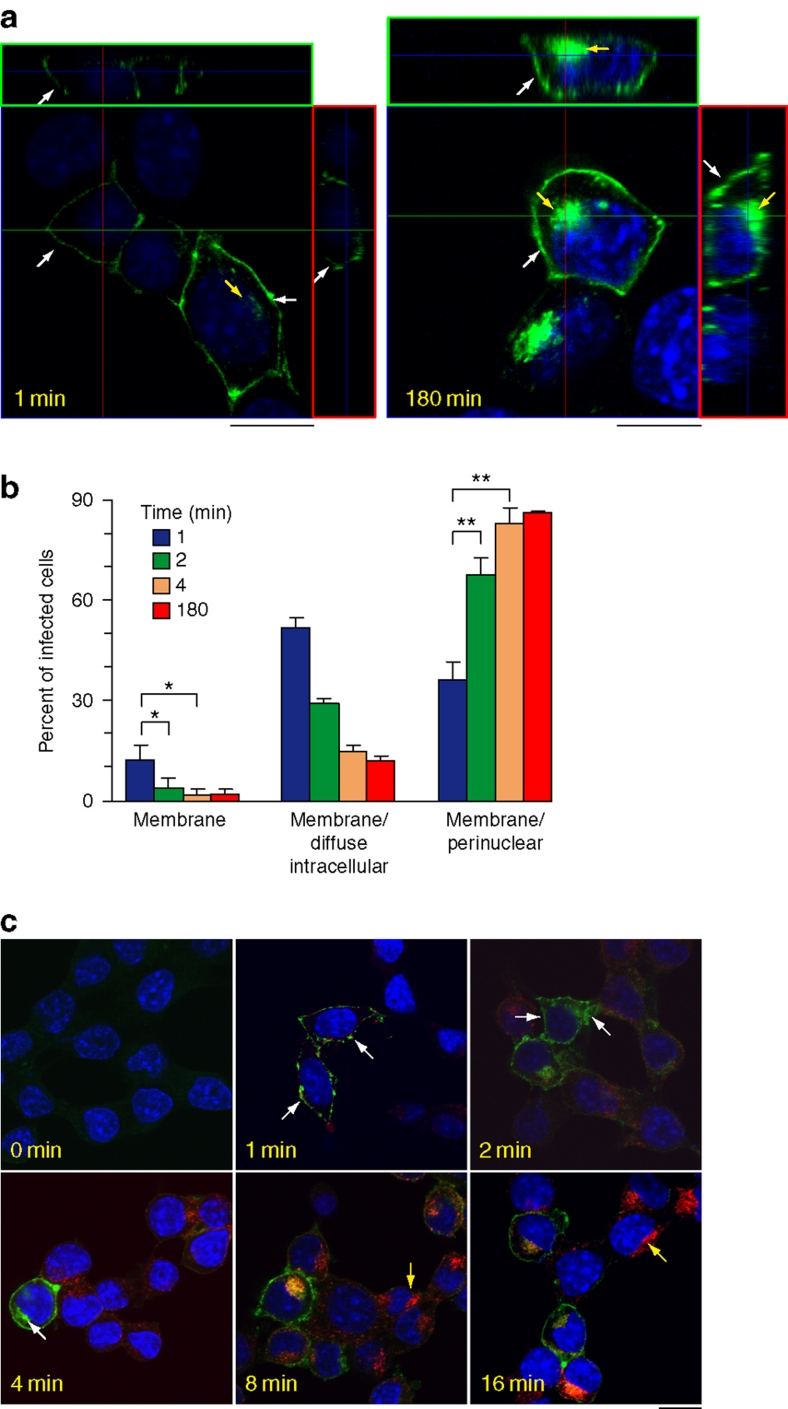
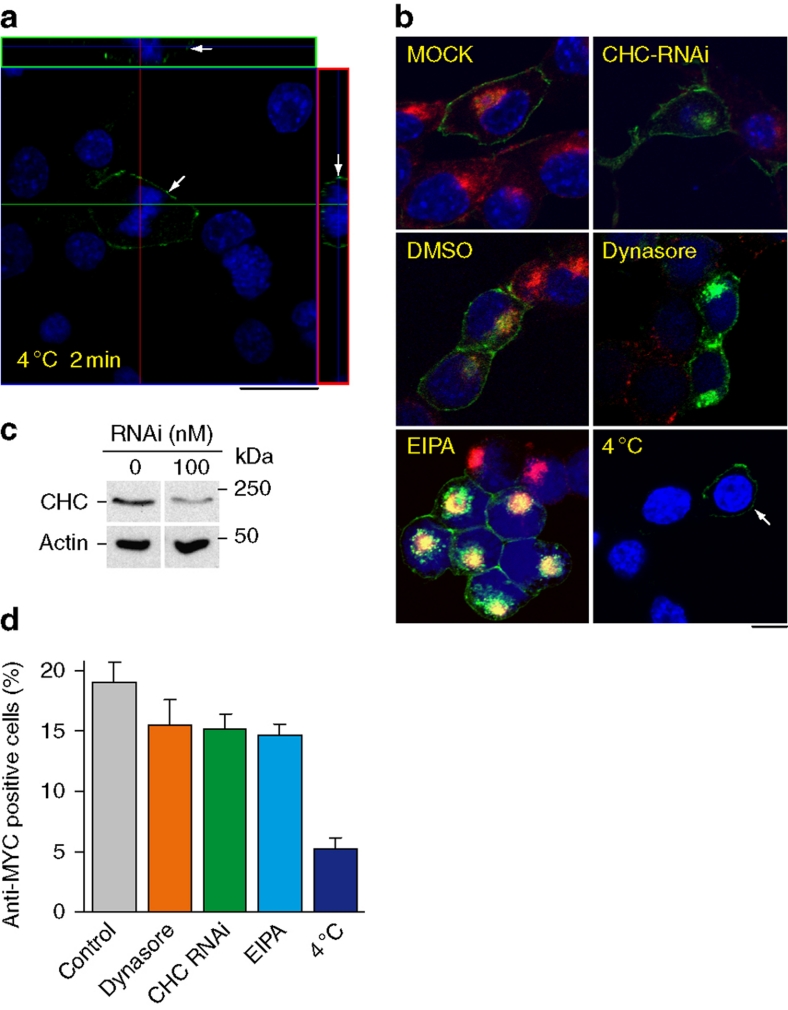
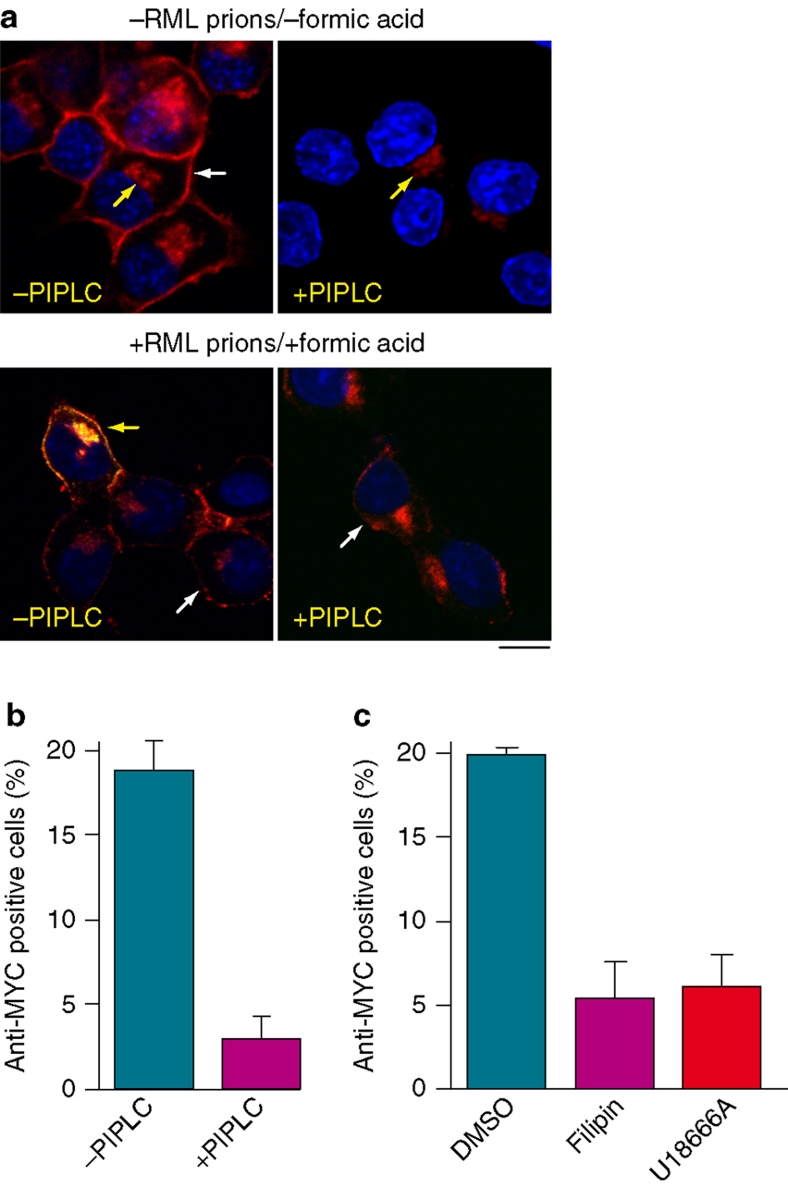
Prion diseases are fatal neurodegenerative disorders with unique transmissible properties. The infectious and pathological agent is thought to be a misfolded conformer of the prion protein. Little is known about the initial events in prion infection because the infecting prion source has been immunologically indistinguishable from normal cellular prion protein (PrP(C)). Here we develop a unique cell system in which epitope-tagged PrP(C) is expressed in a PrP knockdown (KD) neuroblastoma cell line. The tagged PrP(C), when expressed in our PrP-KD cells, supports prion replication with the production of bona fide epitope-tagged infectious misfolded PrP (PrP(Sc)). Using this epitope-tagged PrP(Sc), we study the earliest events in cellular prion infection and PrP misfolding. We show that prion infection of cells is extremely rapid occurring within 1 min of prion exposure, and we demonstrate that the plasma membrane is the primary site of prion conversion.
Figures
Comment in
-
Prions: The fast and the furious.Nat Rev Mol Cell Biol. 2011 May;12(5):278. doi: 10.1038/nrm3111. Nat Rev Mol Cell Biol. 2011. PMID: 21508984 No abstract available.
References
-
- Prusiner S. B. Novel proteinaceous infectious particles cause scrapie. Science 216, 136–144 (1982). - PubMed
-
- Collinge J. Prion diseases of humans and animals: their causes and molecular basis. Annu. Rev. Neurosci. 24, 519–550 (2001). - PubMed
-
- Goedert M., Clavaguera F. & Tolnay M. The propagation of prion-like protein inclusions in neurodegenerative diseases. Trends Neurosci. 33, 317–325 (2010). - PubMed
-
- Alais S. et al.. Mouse neuroblastoma cells release prion infectivity associated with exosomal vesicles. Biol. Cell 100, 603–615 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
