

Bone proteins PHEX and DMP1 regulate fibroblastic growth factor Fgf23 expression in osteocytes through a common pathway involving FGF receptor (FGFR) signaling
- PMID: 21507898
- PMCID: PMC3136343
- DOI: 10.1096/fj.10-177816
Bone proteins PHEX and DMP1 regulate fibroblastic growth factor Fgf23 expression in osteocytes through a common pathway involving FGF receptor (FGFR) signaling
Abstract
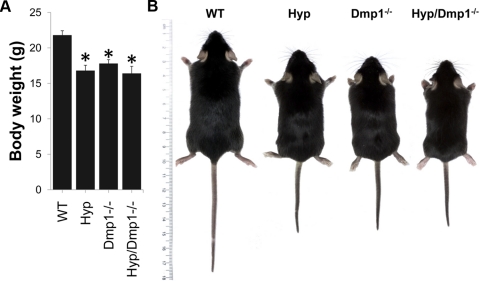
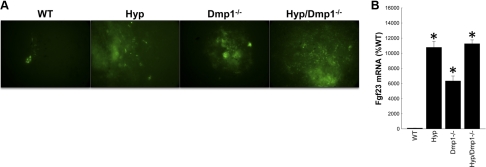
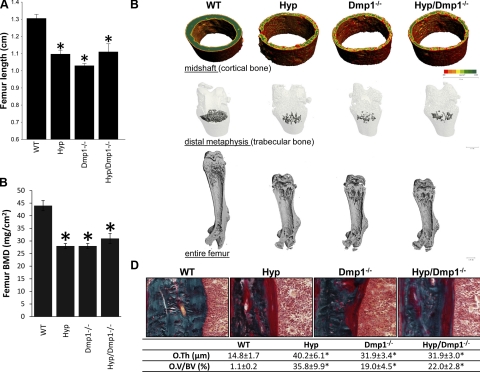
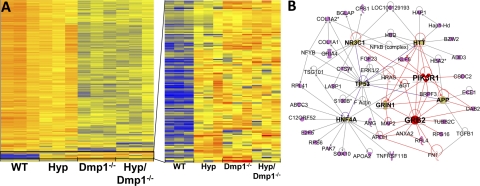
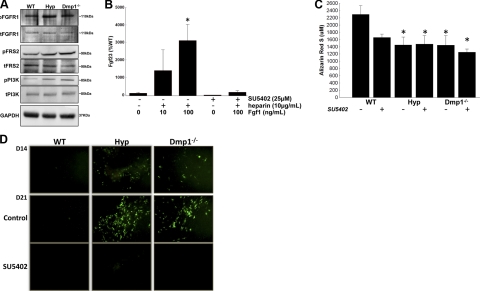
Fibroblastic growth factor 23 (FGF23) is a circulating phosphaturic hormone. Inactivating mutations of the endopeptidase PHEX or the SIBLING protein DMP1 result in equivalent intrinsic bone mineralization defects and increased Fgf23 expression in osteocytes. The mechanisms whereby PHEX and DMP1 regulate Fgf23 expression are unknown. We examined the possibility that PHEX and DMP1 regulate Fgf23 through a common pathway by analyzing the phenotype of compound Phex and Dmp1 mutant mice (Hyp/Dmp1(-/-)). Compared to single-mutant littermates, compound-mutant Hyp/Dmp1(-/-) mice displayed nonadditive elevations of serum FGF23 (1912 ± 183, 1715 ± 178, and 1799 ± 181 pg/ml), hypophosphatemia (P(i): 6.0 ± 0.3, 5.8 ± 0.2, and 5.4 ± 0.1 mg/dl), and severity of rickets/osteomalacia (bone mineral density: -36, -36, and -30%). Microarray analysis of long bones identified gene expression profiles implicating common activation of the FGFR pathway in all the mutant groups. Furthermore, inhibiting FGFR signaling using SU5402 in Hyp- and Dmp1(-/-)-derived bone marrow stromal cells prevented the increase in Fgf23 mRNA expression (129- and 124-fold increase in Hyp and Dmp1(-/-) vs. 1.3-fold in Hyp+SU5402 and 2.5-fold in Dmp1(-/-)+SU5402, P<0.05). For all analyses, samples collected from nonmutant wild-type littermates served as controls. These findings indicate that PHEX and DMP1 control a common pathway regulating bone mineralization and FGF23 production, the latter involving activation of the FGFR signaling in osteocytes.
Figures
References
-
- Itoh N., Ornitz D. M. (2004) Evolution of the Fgf and Fgfr gene families. Trends Genet. 20, 563–569 - PubMed
-
- Yamashita T., Yoshioka M., Itoh N. (2000) Identification of a novel fibroblast growth factor, FGF-23, preferentially expressed in the ventrolateral thalamic nucleus of the brain. Biochem. Biophys. Res. Commun. 277, 494–498 - PubMed
-
- Urakawa I., Yamazaki Y., Shimada T., Iijima K., Hasegawa H., Okawa K., Fujita T., Fukumoto S., Yamashita T. (2006) Klotho converts canonical FGF receptor into a specific receptor for FGF23. Nature 444, 770–774
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
