

Polo-like kinase1 is required for recruitment of dynein to kinetochores during mitosis
- PMID: 21507953
- PMCID: PMC3121523
- DOI: 10.1074/jbc.M111.226605
Polo-like kinase1 is required for recruitment of dynein to kinetochores during mitosis
Abstract
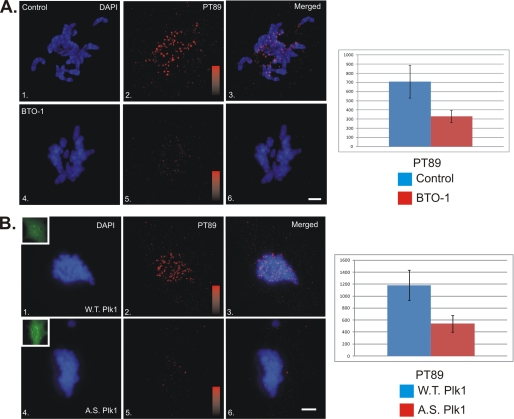
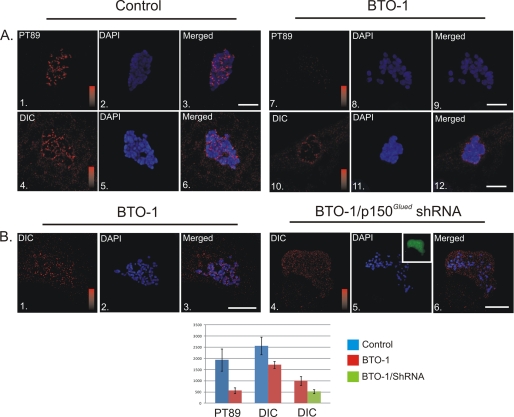
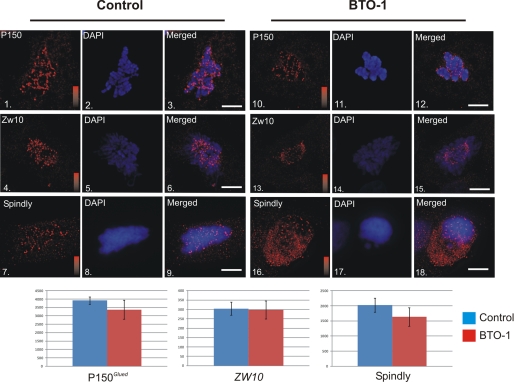
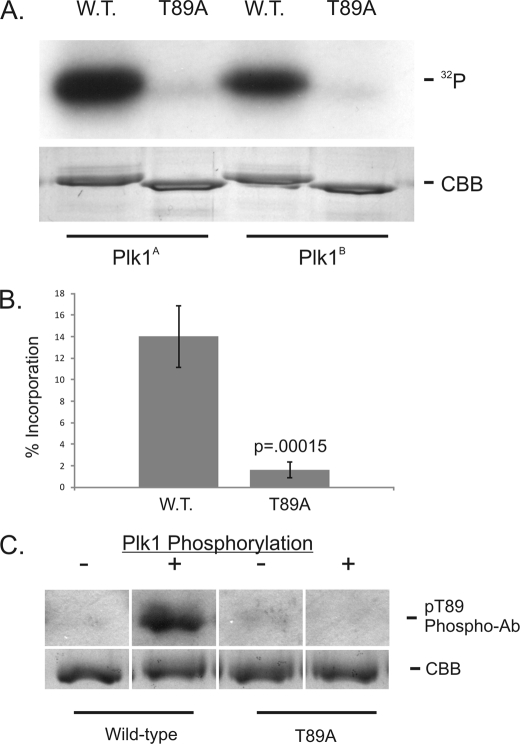
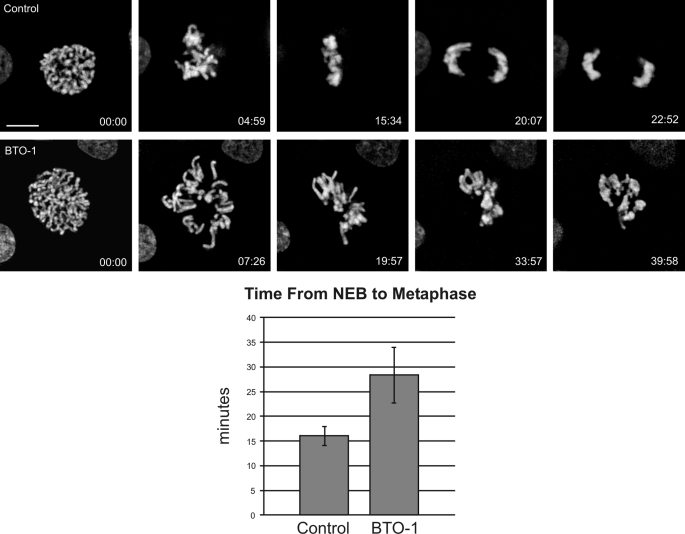
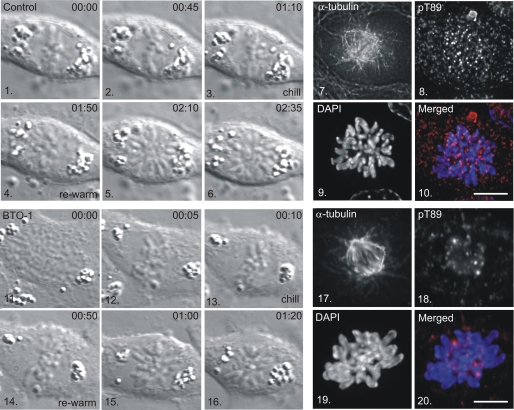
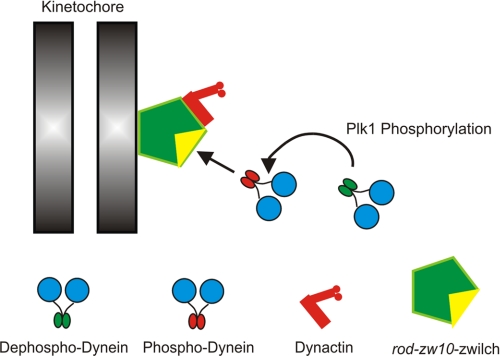
Kinetochore dynein has been implicated in microtubule capture, correcting inappropriate microtubule attachments, chromosome movement, and checkpoint silencing. It remains unclear how dynein coordinates this diverse set of functions. Phosphorylation is responsible for some dynein heterogeneity (Whyte, J., Bader, J. R., Tauhata, S. B., Raycroft, M., Hornick, J., Pfister, K. K., Lane, W. S., Chan, G. K., Hinchcliffe, E. H., Vaughan, P. S., and Vaughan, K. T. (2008) J. Cell Biol. 183, 819-834), and phosphorylated and dephosphorylated forms of dynein coexist at prometaphase kinetochores. In this study, we measured the impact of inhibiting polo-like kinase 1 (Plk1) on both dynein populations. Phosphorylated dynein was ablated at kinetochores after inhibiting Plk1 with a small molecule inhibitor (5-Cyano-7-nitro-2-(benzothiazolo-N-oxide)-carboxamide) or chemical genetic approaches. The total complement of kinetochore dynein was also reduced but not eliminated, reflecting the presence of some dephosphorylated dynein after Plk1 inhibition. Although Plk1 inhibition had a profound effect on dynein, kinetochore populations of dynactin, spindly, and zw10 were not reduced. Plk1-independent dynein was reduced after p150(Glued) depletion, consistent with the binding of dephosphorylated dynein to dynactin. Plk1 phosphorylated dynein intermediate chains at Thr-89 in vitro and generated the phospho-Thr-89 phospho-epitope on recombinant dynein intermediate chains. Finally, inhibition of Plk1 induced defects in microtubule capture and persistent microtubule attachment, suggesting a role for phosphorylated dynein in these functions during prometaphase. These findings suggest that Plk1 is a dynein kinase required for recruitment of phosphorylated dynein to kinetochores.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
