

Covalent structural changes in unfolded GroES that lead to amyloid fibril formation detected by NMR: insight into intrinsically disordered proteins
- PMID: 21507961
- PMCID: PMC3122234
- DOI: 10.1074/jbc.M111.228445
Covalent structural changes in unfolded GroES that lead to amyloid fibril formation detected by NMR: insight into intrinsically disordered proteins
Abstract
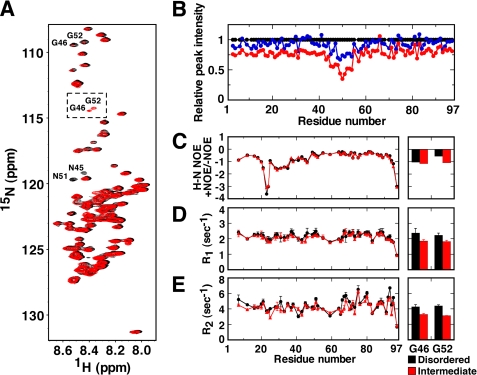
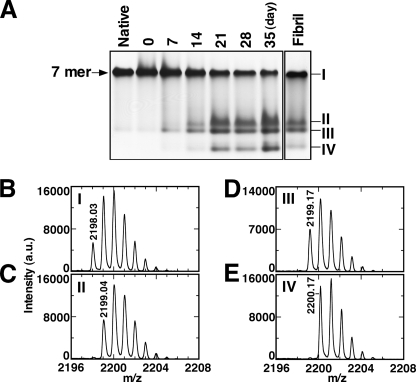
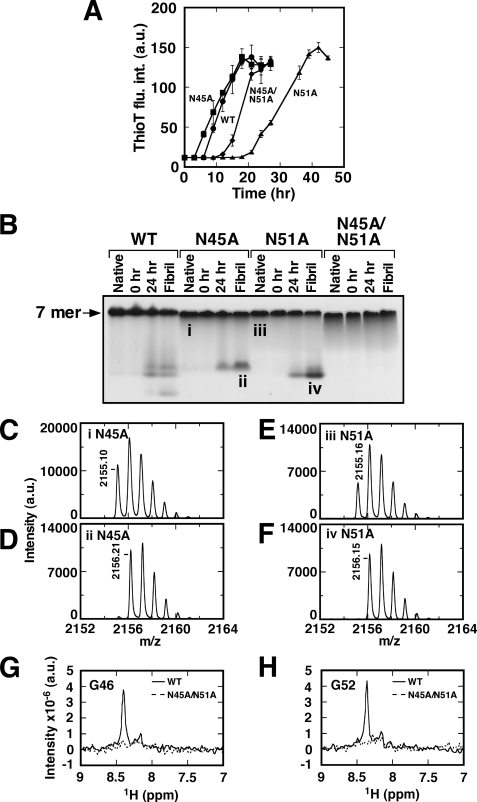
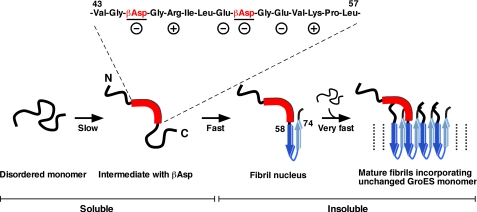
Co-chaperonin GroES from Escherichia coli works with chaperonin GroEL to mediate the folding reactions of various proteins. However, under specific conditions, i.e. the completely disordered state in guanidine hydrochloride, this molecular chaperone forms amyloid fibrils similar to those observed in various neurodegenerative diseases. Thus, this is a good model system to understand the amyloid fibril formation mechanism of intrinsically disordered proteins. Here, we identified a critical intermediate of GroES in the early stages of this fibril formation using NMR and mass spectroscopy measurements. A covalent rearrangement of the polypeptide bond at Asn(45)-Gly(46) and/or Asn(51)-Gly(52) that eventually yield β-aspartic acids via deamidation of asparagine was observed to precede fibril formation. Mutation of these asparagines to alanines resulted in delayed nucleus formation. Our results indicate that peptide bond rearrangement at Asn-Gly enhances the formation of GroES amyloid fibrils. The finding provides a novel insight into the structural process of amyloid fibril formation from a disordered state, which may be applicable to intrinsically disordered proteins in general.
Figures


Similar articles
-
Fibril formation of hsp10 homologue proteins and determination of fibril core regions: differences in fibril core regions dependent on subtle differences in amino acid sequence.J Mol Biol. 2008 Apr 11;377(5):1593-606. doi: 10.1016/j.jmb.2008.02.012. Epub 2008 Feb 14. J Mol Biol. 2008. PMID: 18329043
-
Amyloid-like fibril formation of co-chaperonin GroES: nucleation and extension prefer different degrees of molecular compactness.J Mol Biol. 2005 Sep 2;351(5):1057-69. doi: 10.1016/j.jmb.2005.07.006. J Mol Biol. 2005. PMID: 16054644
-
Chaperonin GroEL accelerates protofibril formation and decorates fibrils of the Het-s prion protein.Proc Natl Acad Sci U S A. 2017 Aug 22;114(34):9104-9109. doi: 10.1073/pnas.1711645114. Epub 2017 Aug 7. Proc Natl Acad Sci U S A. 2017. PMID: 28784759 Free PMC article.
-
GroEL and the GroEL-GroES Complex.Subcell Biochem. 2017;83:483-504. doi: 10.1007/978-3-319-46503-6_17. Subcell Biochem. 2017. PMID: 28271487 Review.
-
Reaction Cycle of Chaperonin GroEL via Symmetric "Football" Intermediate.J Mol Biol. 2015 Sep 11;427(18):2912-8. doi: 10.1016/j.jmb.2015.04.007. Epub 2015 Apr 18. J Mol Biol. 2015. PMID: 25900372 Review.
Cited by
-
Probing the functional mechanism of Escherichia coli GroEL using circular permutation.PLoS One. 2011;6(10):e26462. doi: 10.1371/journal.pone.0026462. Epub 2011 Oct 18. PLoS One. 2011. PMID: 22028884 Free PMC article.
-
Asparagine deamidation reduces DNA-binding affinity of the Drosophila melanogaster Scr homeodomain.FEBS Lett. 2015 Oct 24;589(21):3237-41. doi: 10.1016/j.febslet.2015.09.020. Epub 2015 Oct 3. FEBS Lett. 2015. PMID: 26435141 Free PMC article.
-
Kinetic, thermodynamic, and ab initio insights of AsnGly isomerisation as a ticking time bomb for protein integrity.Commun Chem. 2024 Dec 19;7(1):303. doi: 10.1038/s42004-024-01374-1. Commun Chem. 2024. PMID: 39702829 Free PMC article.
-
Stability of Protein Pharmaceuticals: Recent Advances.Pharm Res. 2024 Jul;41(7):1301-1367. doi: 10.1007/s11095-024-03726-x. Epub 2024 Jun 27. Pharm Res. 2024. PMID: 38937372 Review.
-
Suppression of amyloid fibrils using the GroEL apical domain.Sci Rep. 2016 Aug 4;6:31041. doi: 10.1038/srep31041. Sci Rep. 2016. PMID: 27488469 Free PMC article.
References
-
- Wright P. E., Dyson H. J. (1999) J. Mol. Biol. 293, 321–331 - PubMed
-
- Sugase K., Dyson H. J., Wright P. E. (2007) Nature 447, 1021–1025 - PubMed
-
- Ward J. J., Sodhi J. S., McGuffin L. J., Buxton B. F., Jones D. T. (2004) J. Mol. Biol. 337, 635–645 - PubMed
-
- Spillantini M. G., Schmidt M. L., Lee V. M., Trojanowski J. Q., Jakes R., Goedert M. (1997) Nature 388, 839–840 - PubMed
-
- Yagi H., Kusaka E., Hongo K., Mizobata T., Kawata Y. (2005) J. Biol. Chem. 280, 38609–38616 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
