

The SUMOylation of matrix protein M1 modulates the assembly and morphogenesis of influenza A virus
- PMID: 21507966
- PMCID: PMC3126537
- DOI: 10.1128/JVI.02401-10
The SUMOylation of matrix protein M1 modulates the assembly and morphogenesis of influenza A virus
Abstract
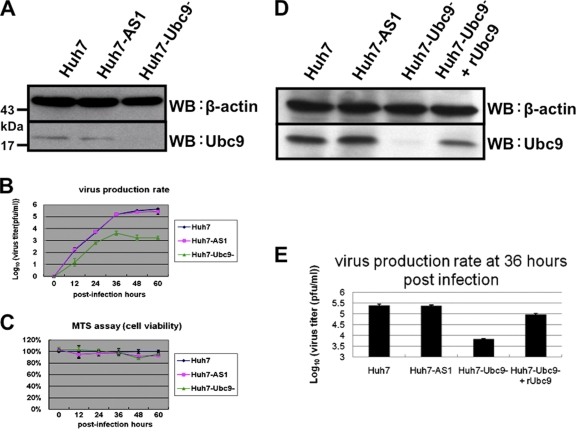
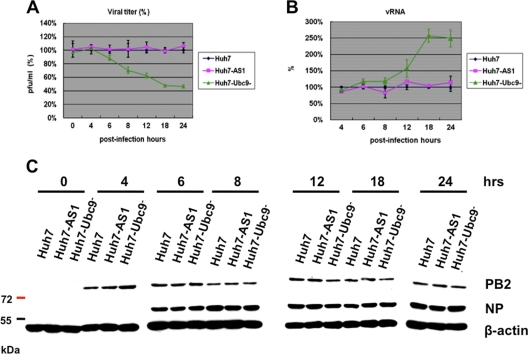
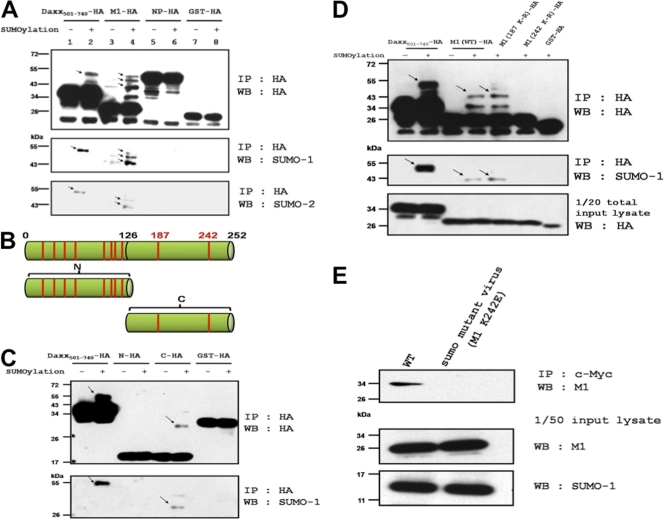
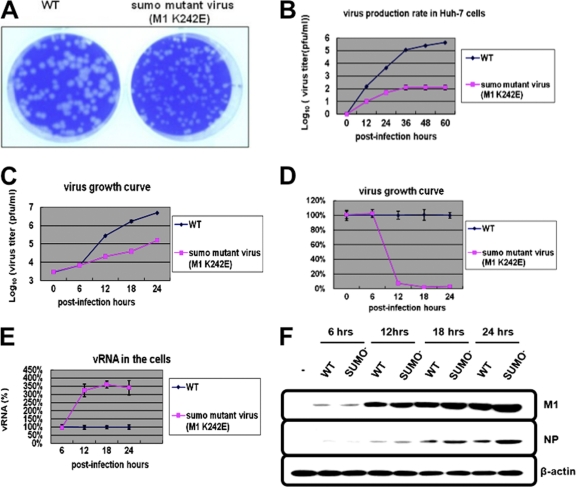
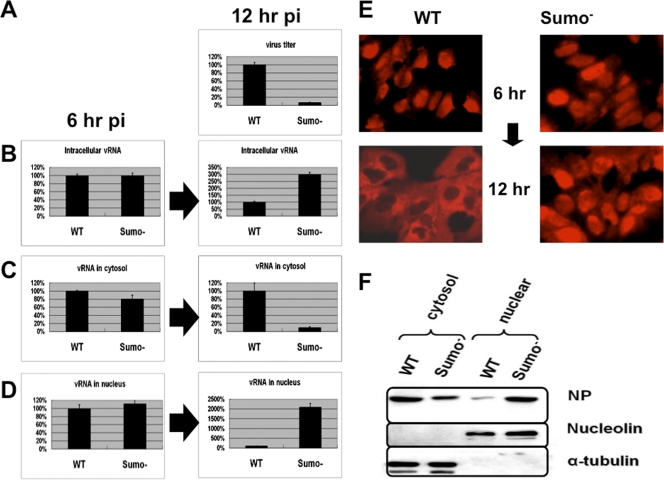
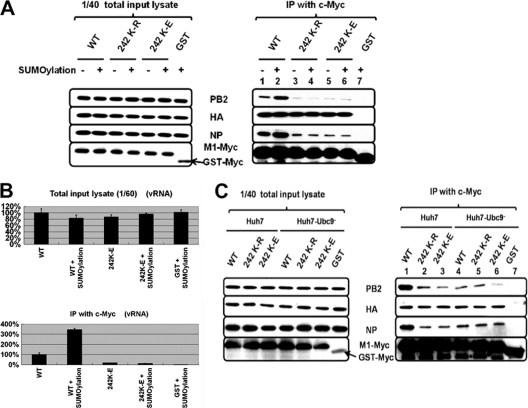
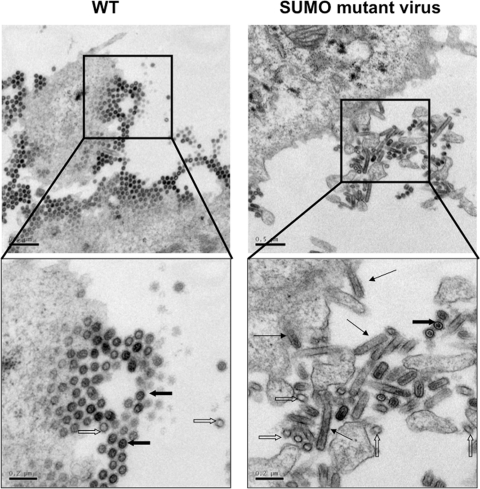
SUMOylation is an important posttranslational modification for regulation of cellular functions and viral replication. Here, we report that protein SUMOylation regulates the replication of influenza A virus at the steps of viral maturation and assembly. Knocking down the SUMO-conjugating enzyme Ubc9 resulted in the reduction of virus production. Dissection of the virus life cycle revealed that SUMOylation is involved in the processes of virus maturation and assembly. The viral matrix protein M1 is SUMOylated at K242. A virus carrying the SUMO-defective M1 produced a lower titer of virus, while its viral proteins and viral RNA (vRNA) accumulated in the cells. Furthermore, the mechanistic studies showed that the SUMOylation of M1 is required for the interaction between M1 and viral RNP (vRNP) to form the M1-vRNP complex. The lack of M1 SUMOylation prevented the nuclear export of vRNP and subsequent viral morphogenesis. Taken together, our findings elucidate that the maturation and assembly of influenza A virus is controlled by the SUMO modification of M1 protein. Therefore, we suggest that M1 can serve as a target for developing a new generation of drugs for flu therapy.
Figures
Similar articles
-
Nuclear export of influenza viral ribonucleoprotein is temperature-dependently inhibited by dissociation of viral matrix protein.Virology. 2003 Feb 15;306(2):244-53. doi: 10.1016/s0042-6822(02)00013-2. Virology. 2003. PMID: 12642098
-
Selective incorporation of vRNP into influenza A virions determined by its specific interaction with M1 protein.Virology. 2017 May;505:23-32. doi: 10.1016/j.virol.2017.02.008. Epub 2017 Feb 17. Virology. 2017. PMID: 28219018 Free PMC article.
-
Interaction of NS2 with AIMP2 facilitates the switch from ubiquitination to SUMOylation of M1 in influenza A virus-infected cells.J Virol. 2015 Jan;89(1):300-11. doi: 10.1128/JVI.02170-14. Epub 2014 Oct 15. J Virol. 2015. PMID: 25320310 Free PMC article.
-
Assembly and budding of influenza virus.Virus Res. 2004 Dec;106(2):147-65. doi: 10.1016/j.virusres.2004.08.012. Virus Res. 2004. PMID: 15567494 Free PMC article. Review.
-
[Structure, function and regulation of expression of influenza virus matrix M1 protein].Nihon Rinsho. 1997 Oct;55(10):2581-6. Nihon Rinsho. 1997. PMID: 9360375 Review. Japanese.
Cited by
-
Broad-spectrum antiviral agents.Front Microbiol. 2015 May 22;6:517. doi: 10.3389/fmicb.2015.00517. eCollection 2015. Front Microbiol. 2015. PMID: 26052325 Free PMC article. Review.
-
Thapsigargin at Non-Cytotoxic Levels Induces a Potent Host Antiviral Response that Blocks Influenza A Virus Replication.Viruses. 2020 Sep 27;12(10):1093. doi: 10.3390/v12101093. Viruses. 2020. PMID: 32992478 Free PMC article.
-
PIAS1-mediated SUMOylation of influenza A virus PB2 restricts viral replication and virulence.PLoS Pathog. 2022 Apr 4;18(4):e1010446. doi: 10.1371/journal.ppat.1010446. eCollection 2022 Apr. PLoS Pathog. 2022. PMID: 35377920 Free PMC article.
-
NP and NS1 proteins of H5N1 virus significantly upregulated IFITM1, IFITM2, and IFITM3 in A549 cells.Afr Health Sci. 2019 Mar;19(1):1402-1410. doi: 10.4314/ahs.v19i1.13. Afr Health Sci. 2019. PMID: 31148967 Free PMC article.
-
Influenza virus-host interactome screen as a platform for antiviral drug development.Cell Host Microbe. 2014 Dec 10;16(6):795-805. doi: 10.1016/j.chom.2014.11.002. Epub 2014 Nov 20. Cell Host Microbe. 2014. PMID: 25464832 Free PMC article.
References
-
- Arzt S., Petit I., Burmeister W. P., Ruigrok R. W., Baudin F. 2004. Structure of a knockout mutant of influenza virus M1 protein that has altered activities in membrane binding, oligomerisation and binding to NEP (NS2). Virus Res. 99:115–119 - PubMed
-
- Baudin F., Petit I., Weissenhorn W., Ruigrok R. W. 2001. In vitro dissection of the membrane and RNP binding activities of influenza virus M1 protein. Virology 281:102–108 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
