

RNA splicing in a new rhabdovirus from Culex mosquitoes
- PMID: 21507977
- PMCID: PMC3126488
- DOI: 10.1128/JVI.00040-11
RNA splicing in a new rhabdovirus from Culex mosquitoes
Abstract
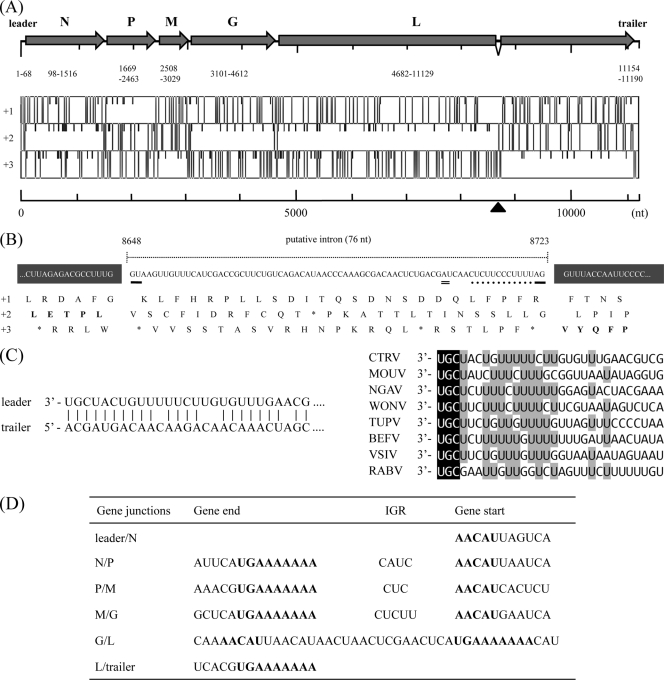
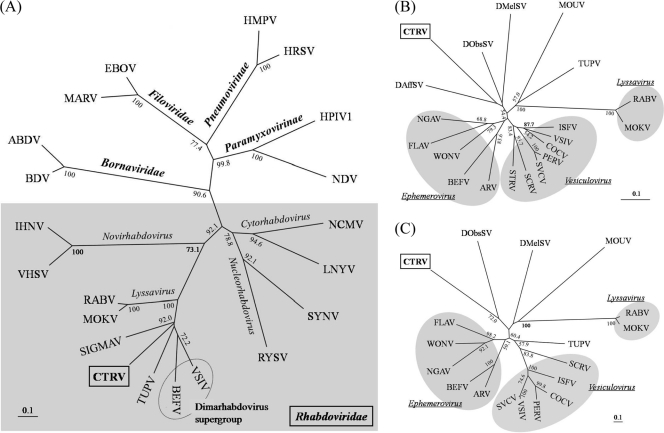
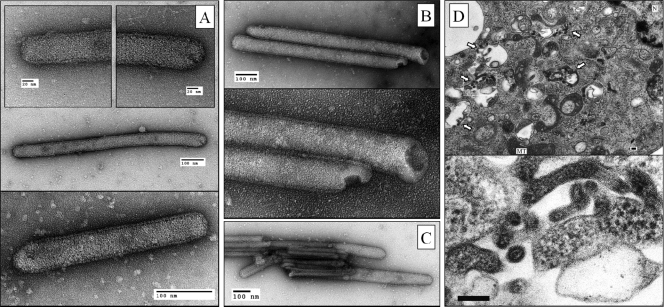
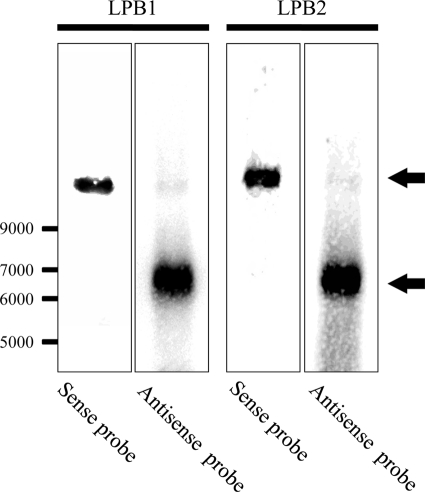
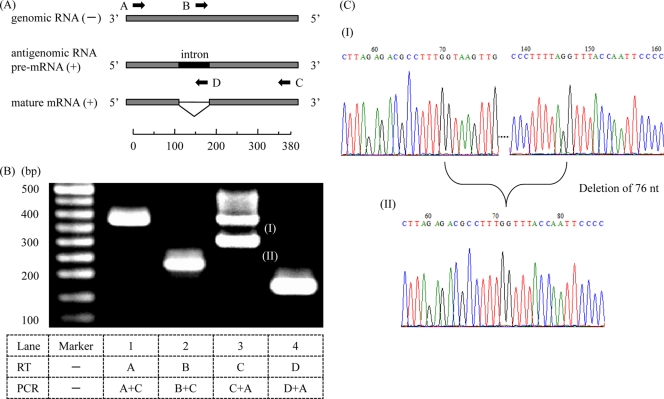
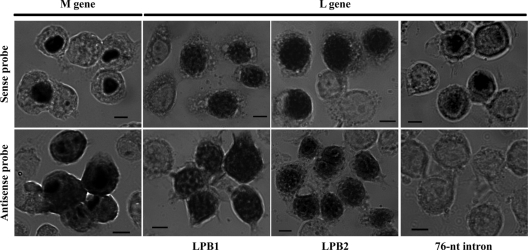
Among members of the order Mononegavirales, RNA splicing events have been found only in the family Bornaviridae. Here, we report that a new rhabdovirus isolated from the mosquito Culex tritaeniorhynchus replicates in the nuclei of infected cells and requires RNA splicing for viral mRNA maturation. The virus, designated Culex tritaeniorhynchus rhabdovirus (CTRV), shares a similar genome organization with other rhabdoviruses, except for the presence of a putative intron in the coding region for the L protein. Molecular phylogenetic studies indicated that CTRV belongs to the family Rhabdoviridae, but it is yet to be assigned a genus. Electron microscopic analysis revealed that the CTRV virion is extremely elongated, unlike virions of rhabdoviruses, which are generally bullet shaped. Northern hybridization confirmed that a large transcript (approximately 6,500 nucleotides [nt]) from the CTRV L gene was present in the infected cells. Strand-specific reverse transcription-PCR (RT-PCR) analyses identified the intron-exon boundaries and the 76-nt intron sequence, which contains the typical motif for eukaryotic spliceosomal intron-splice donor/acceptor sites (GU-AG), a predicted branch point, and a polypyrimidine tract. In situ hybridization exhibited that viral RNAs are primarily localized in the nucleus of infected cells, indicating that CTRV replicates in the nucleus and is allowed to utilize the host's nuclear splicing machinery. This is the first report of RNA splicing among the members of the family Rhabdoviridae.
Figures
Similar articles
-
Persistent natural infection of a Culex tritaeniorhynchus cell line with a novel Culex tritaeniorhynchus rhabdovirus strain.Microbiol Immunol. 2015 Sep;59(9):562-6. doi: 10.1111/1348-0421.12279. Microbiol Immunol. 2015. PMID: 26112738
-
Characterisation of a novel nucleorhabdovirus infecting alfalfa (Medicago sativa).Virol J. 2019 Apr 29;16(1):55. doi: 10.1186/s12985-019-1147-3. Virol J. 2019. PMID: 31036009 Free PMC article.
-
Moussa virus: a new member of the Rhabdoviridae family isolated from Culex decens mosquitoes in Côte d'Ivoire.Virus Res. 2010 Jan;147(1):17-24. doi: 10.1016/j.virusres.2009.09.013. Epub 2009 Oct 3. Virus Res. 2010. PMID: 19804801 Free PMC article.
-
[Plant rhabdoviruses with bipartite genomes].Uirusu. 2013;63(2):143-54. doi: 10.2222/jsv.63.143. Uirusu. 2013. PMID: 25366049 Review. Japanese.
-
Structural insights into the rhabdovirus transcription/replication complex.Virus Res. 2011 Dec;162(1-2):126-37. doi: 10.1016/j.virusres.2011.09.025. Epub 2011 Sep 22. Virus Res. 2011. PMID: 21963663 Review.
Cited by
-
Discovery of Culex pipiens associated tunisia virus: a new ssRNA(+) virus representing a new insect associated virus family.Virus Evol. 2018 Jan 10;4(1):vex040. doi: 10.1093/ve/vex040. eCollection 2018 Jan. Virus Evol. 2018. PMID: 29340209 Free PMC article.
-
Novel viruses in salivary glands of mosquitoes from sylvatic Cerrado, Midwestern Brazil.PLoS One. 2017 Nov 8;12(11):e0187429. doi: 10.1371/journal.pone.0187429. eCollection 2017. PLoS One. 2017. PMID: 29117239 Free PMC article.
-
Identification of a novel rhabdovirus in Spodoptera frugiperda cell lines.J Virol. 2014 Jun;88(12):6576-85. doi: 10.1128/JVI.00780-14. Epub 2014 Mar 26. J Virol. 2014. PMID: 24672045 Free PMC article.
-
Insect-Specific Virus Discovery: Significance for the Arbovirus Community.Viruses. 2015 Sep 10;7(9):4911-28. doi: 10.3390/v7092851. Viruses. 2015. PMID: 26378568 Free PMC article. Review.
-
Metagenomic analysis evidences a core virome in Anopheles darlingi from three contrasting Colombian ecoregions.PLoS One. 2025 Apr 30;20(4):e0320593. doi: 10.1371/journal.pone.0320593. eCollection 2025. PLoS One. 2025. PMID: 40305569 Free PMC article.
References
-
- Ammar el-D., Tsai C. W., Whitfield A. E., Redinbaugh M. G., Hogenhout S. A. 2009. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annu. Rev. Entomol. 54:447–468 - PubMed
-
- Bock J. O., Lundsgaard T., Pedersen P. A., Christensen L. S. 2004. Identification and partial characterization of Taastrup virus: a newly identified member species of the Mononegavirales. Virology 319:49–59 - PubMed
-
- Boulo S., Akarsu H., Ruigrok R. W., Baudin F. 2007. Nuclear traffic of influenza virus proteins and ribonucleoprotein complexes. Virus Res. 124:12–21 - PubMed
-
- Bourhy H., Cowley J. A., Larrous F., Holmes E. C., Walker P. J. 2005. Phylogenetic relationships among rhabdoviruses inferred using the L polymerase gene. J. Gen. Virol. 86:2849–2858 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
