

An analogy between the evolution of drug resistance in bacterial communities and malignant tissues
- PMID: 21508974
- PMCID: PMC3488437
- DOI: 10.1038/nrc3039
An analogy between the evolution of drug resistance in bacterial communities and malignant tissues
Abstract
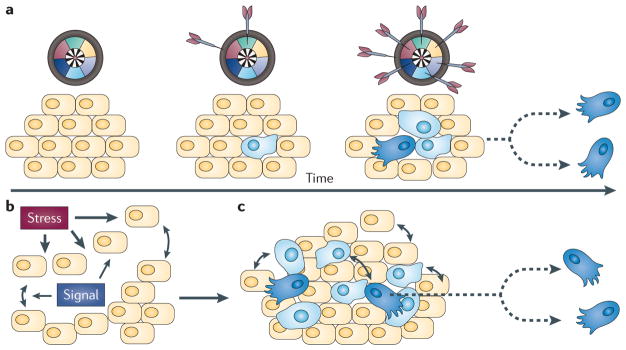
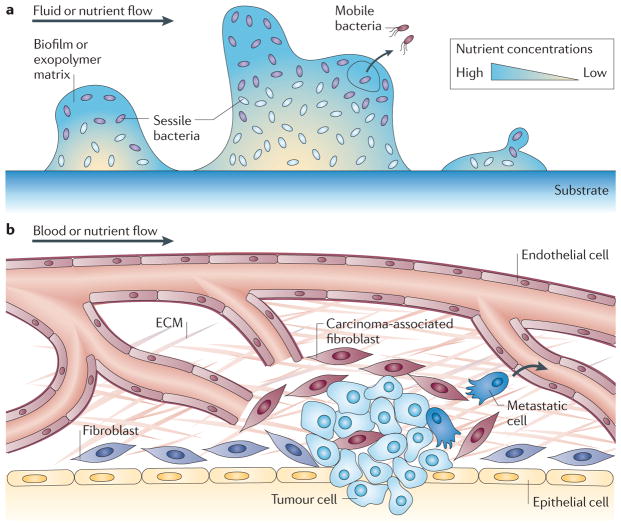
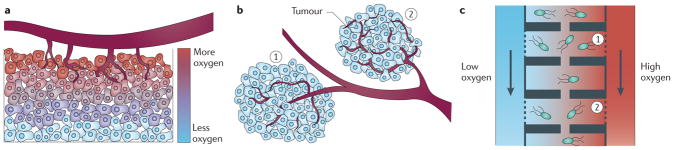
Cancer cells rapidly evolve drug resistance through somatic evolution and, in order to continue growth in the metastatic phase, violate the organism-wide consensus of regulated growth and beneficial communal interactions. We suggest that there is a fundamental mechanistic connection between the rapid evolution of resistance to chemotherapy in cellular communities within malignant tissues and the rapid evolution of antibiotic resistance in bacterial communities. We propose that this evolution is the result of a programmed and collective stress response performed by interacting cells, and that, given this fundamental connection, studying bacterial communities can provide deeper insights into the dynamics of adaptation and the evolution of cells within tumours.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Similar articles
-
On the rapidity of antibiotic resistance evolution facilitated by a concentration gradient.Proc Natl Acad Sci U S A. 2012 Jul 3;109(27):10775-80. doi: 10.1073/pnas.1117716109. Epub 2012 Jun 18. Proc Natl Acad Sci U S A. 2012. PMID: 22711808 Free PMC article.
-
Conflicting objectives in chemotherapy with drug resistance.Bull Math Biol. 1997 Jul;59(4):707-24. doi: 10.1007/BF02458426. Bull Math Biol. 1997. PMID: 9214850
-
Modeling drug resistance in a conjoint normal-tumor setting.Theor Biol Med Model. 2015 Jan 15;12:3. doi: 10.1186/1742-4682-12-3. Theor Biol Med Model. 2015. PMID: 25588472 Free PMC article.
-
Chemotherapy curable malignancies and cancer stem cells: a biological review and hypothesis.BMC Cancer. 2016 Nov 21;16(1):906. doi: 10.1186/s12885-016-2956-z. BMC Cancer. 2016. PMID: 27871274 Free PMC article. Review.
-
Managing drug resistance in cancer: lessons from HIV therapy.Nat Rev Cancer. 2012 Jun 7;12(7):494-501. doi: 10.1038/nrc3297. Nat Rev Cancer. 2012. PMID: 22673150 Review.
Cited by
-
Evolutionary game theory for physical and biological scientists. II. Population dynamics equations can be associated with interpretations.Interface Focus. 2014 Aug 6;4(4):20140038. doi: 10.1098/rsfs.2014.0038. Interface Focus. 2014. PMID: 25097752 Free PMC article. Review.
-
Emerging and evolving concepts in gene essentiality.Nat Rev Genet. 2018 Jan;19(1):34-49. doi: 10.1038/nrg.2017.74. Epub 2017 Oct 16. Nat Rev Genet. 2018. PMID: 29033457 Review.
-
α,α-disubstituted β-amino amides eliminate Staphylococcus aureus biofilms by membrane disruption and biomass removal.Biofilm. 2023 Aug 25;6:100151. doi: 10.1016/j.bioflm.2023.100151. eCollection 2023 Dec 15. Biofilm. 2023. PMID: 37662850 Free PMC article.
-
Stress-induced mutagenesis and complex adaptation.Proc Biol Sci. 2014 Oct 7;281(1792):20141025. doi: 10.1098/rspb.2014.1025. Proc Biol Sci. 2014. PMID: 25143032 Free PMC article.
-
Convection shapes the trade-off between antibiotic efficacy and the selection for resistance in spatial gradients.Phys Biol. 2017 Jul 19;14(4):045011. doi: 10.1088/1478-3975/aa7bb3. Phys Biol. 2017. PMID: 28649977 Free PMC article.
References
-
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100:57–70. - PubMed
-
- Ellis LM, Hicklin DJ. VEGF-targeted therapy: mechanisms of anti-tumour activity. Nature Rev Cancer. 2008;8:579–591. - PubMed
-
- Weinstein IB, Joe A. Oncogene addiction. Cancer Res. 2008;68:3077–3080. - PubMed
-
- Letai AG. Diagnosing and exploiting cancer’s addiction to blocks in apoptosis. Nature Rev Cancer. 2008;8:121–132. - PubMed
-
- Kamb A, Wee S, Lengauer C. Why is cancer drug discovery so difficult? Nature Rev Drug Discov. 2007;6:115–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
