

Interleukin-10 signaling in regulatory T cells is required for suppression of Th17 cell-mediated inflammation
- PMID: 21511185
- PMCID: PMC3088485
- DOI: 10.1016/j.immuni.2011.03.018
Interleukin-10 signaling in regulatory T cells is required for suppression of Th17 cell-mediated inflammation
Abstract
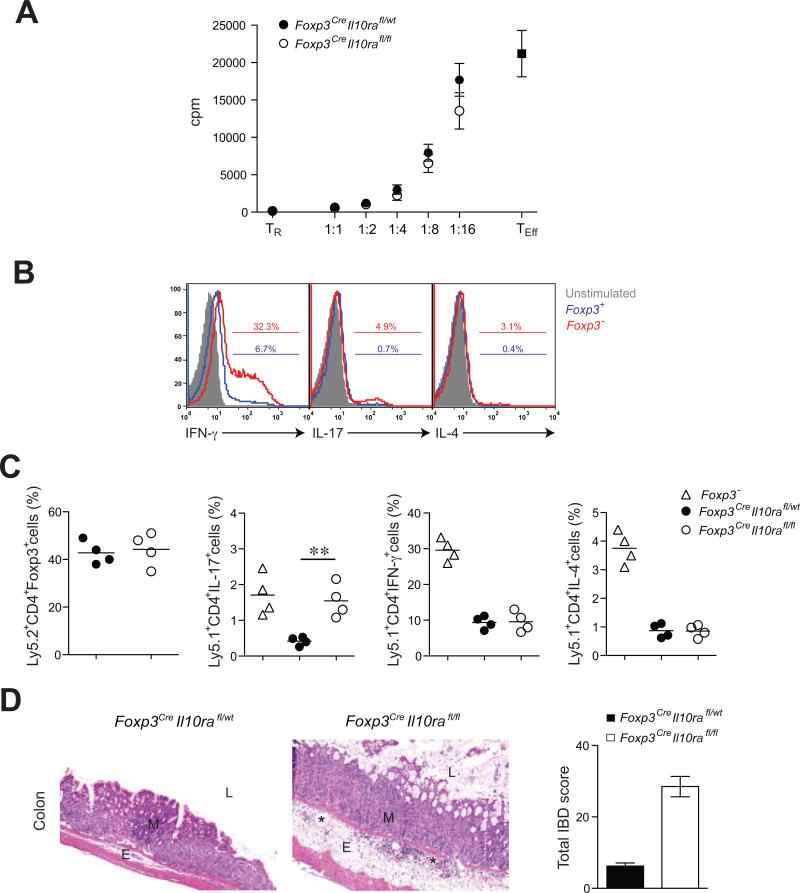
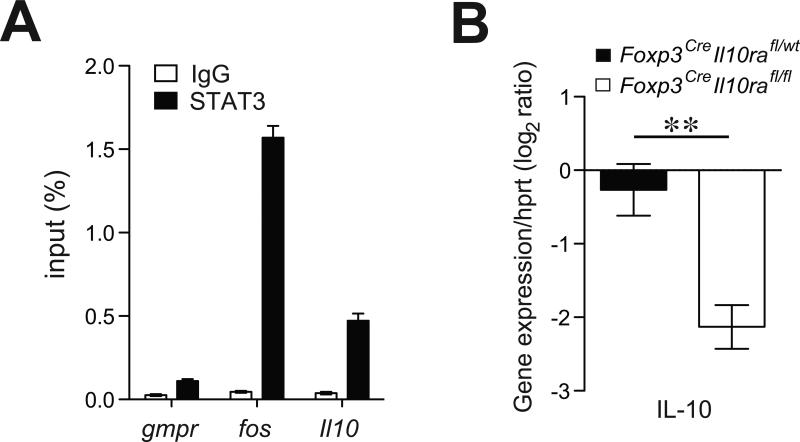
Effector CD4+ T cell subsets, whose differentiation is facilitated by distinct cytokine cues, amplify the corresponding type of inflammatory response. Regulatory T (Treg) cells integrate environmental cues to suppress particular types of inflammation. In this regard, STAT3, a transcription factor essential for T helper 17 (Th17) cell differentiation, is necessary for Treg cell-mediated control of Th17 cell responses. Here, we showed that anti-inflammatory interleukin-10 (IL-10), and not proinflammatory IL-6 and IL-23 cytokine signaling, endowed Treg cells with the ability to suppress pathogenic Th17 cell responses. Ablation of the IL-10 receptor in Treg cells resulted in selective dysregulation of Th17 cell responses and colitis similar to that observed in mice harboring STAT3-deficient Treg cells. Thus, Treg cells limit Th17 cell inflammation by serving as principal amplifiers of negative regulatory circuits operating in immune effector cells.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
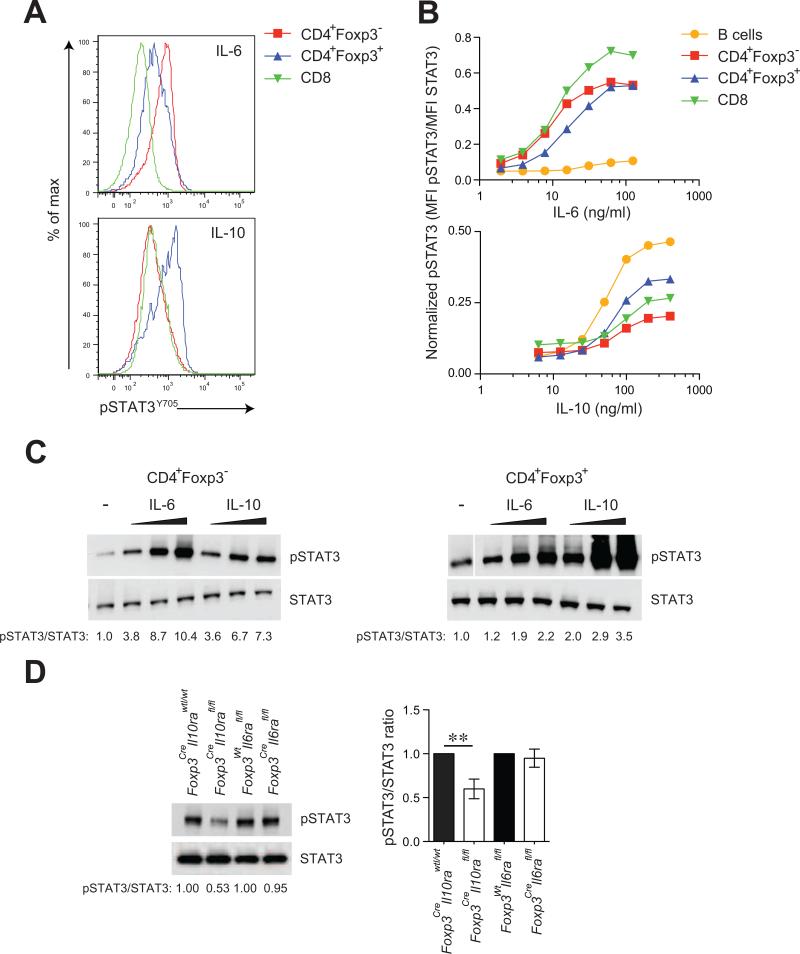
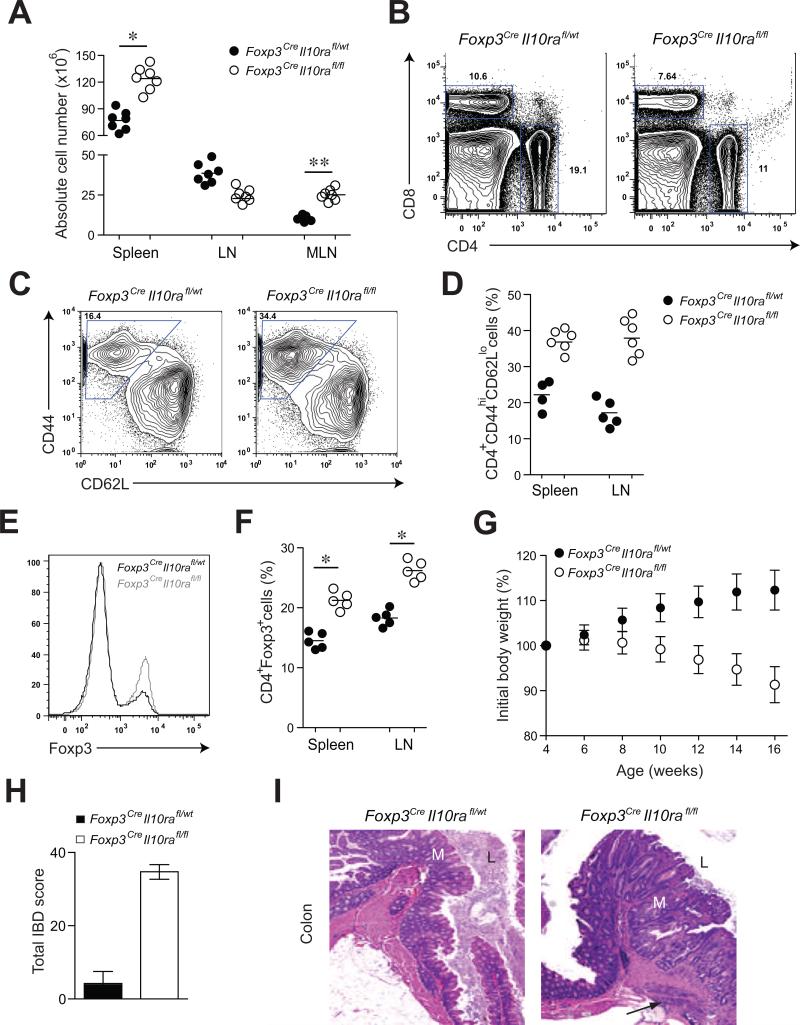
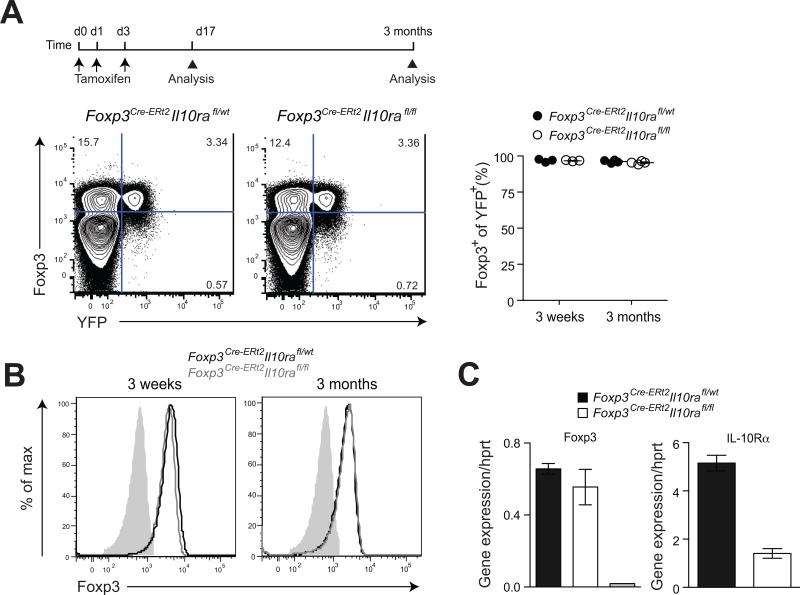
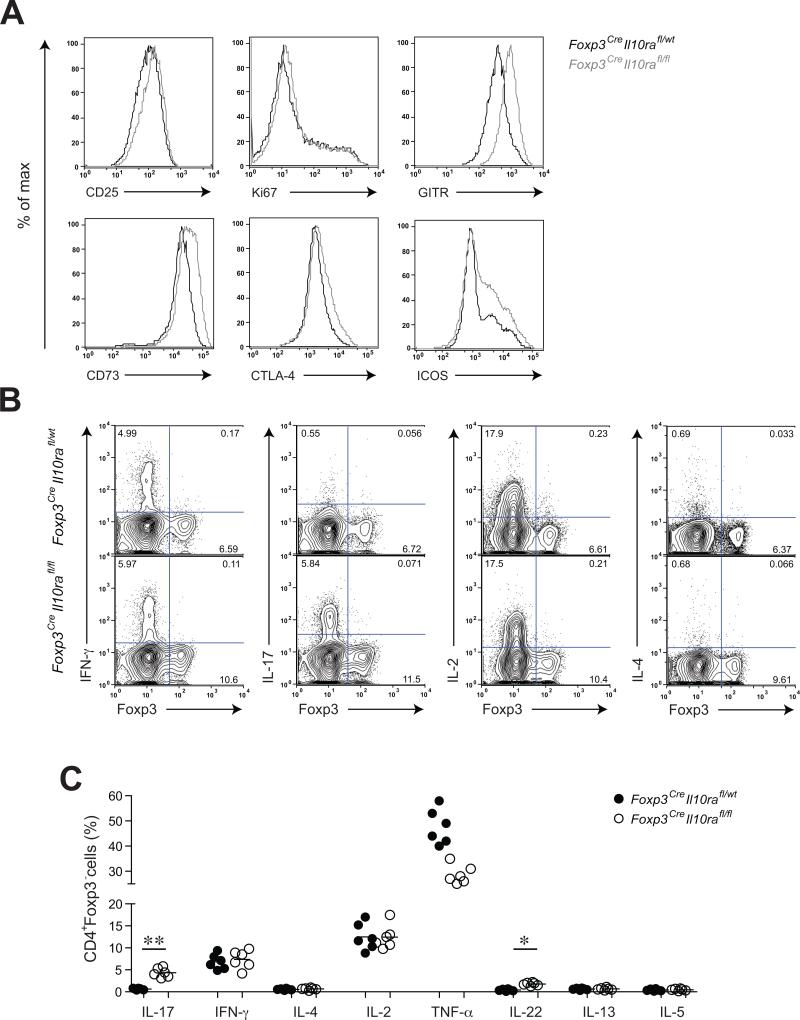
Comment in
-
At 17, in-10's passion need not inflame.Immunity. 2011 Apr 22;34(4):460-2. doi: 10.1016/j.immuni.2011.04.004. Immunity. 2011. PMID: 21511180
References
-
- Ahern PP, Izcue A, Maloy KJ, Powrie F. The interleukin-23 axis in intestinal inflammation. Immunol Rev. 2008;226:147–159. - PubMed
-
- Brand S. Crohn's disease: Th1, Th17 or both? The change of a paradigm: new immunological and genetic insights implicate Th17 cells in the pathogenesis of Crohn's disease. Gut. 2009;58:1152–1167. - PubMed
-
- Burich A, Hershberg R, Waggie K, Zeng W, Brabb T, Westrich G, Viney JL, Maggio-Price L. Helicobacter-induced inflammatory bowel disease in IL-10- and T cell-deficient mice. Am J Physiol Gastrointest Liver Physiol. 2001;281:G764–778. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
