

Computational approaches to RNA structure prediction, analysis, and design
- PMID: 21514143
- PMCID: PMC3112238
- DOI: 10.1016/j.sbi.2011.03.015
Computational approaches to RNA structure prediction, analysis, and design
Abstract
RNA molecules are important cellular components involved in many fundamental biological processes. Understanding the mechanisms behind their functions requires RNA tertiary structure knowledge. Although modeling approaches for the study of RNA structures and dynamics lag behind efforts in protein folding, much progress has been achieved in the past two years. Here, we review recent advances in RNA folding algorithms, RNA tertiary motif discovery, applications of graph theory approaches to RNA structure and function, and in silico generation of RNA sequence pools for aptamer design. Advances within each area can be combined to impact many problems in RNA structure and function.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
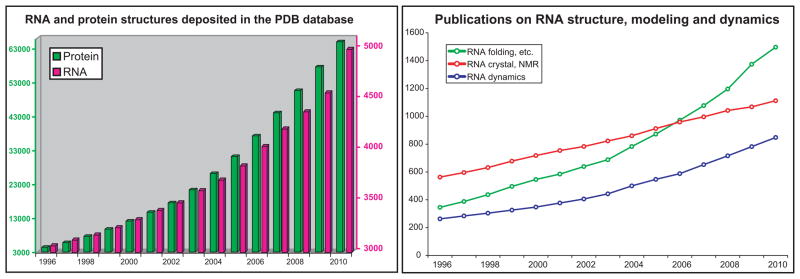
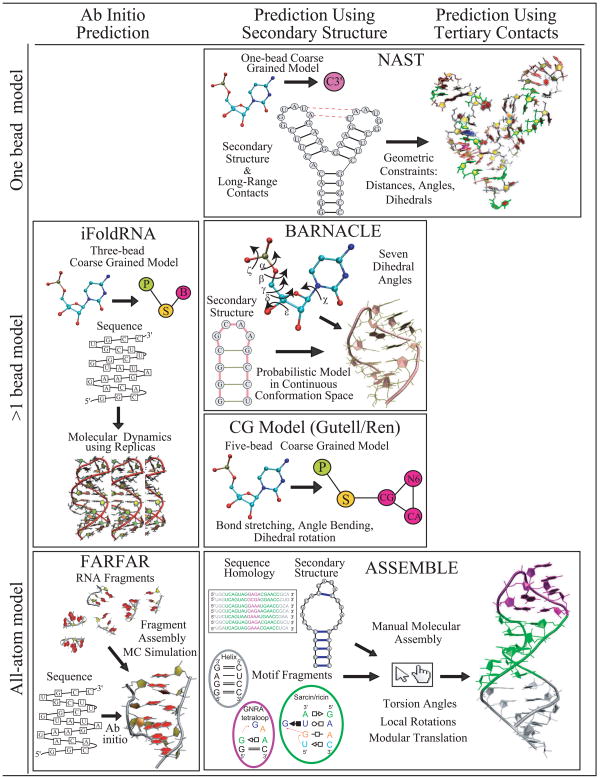
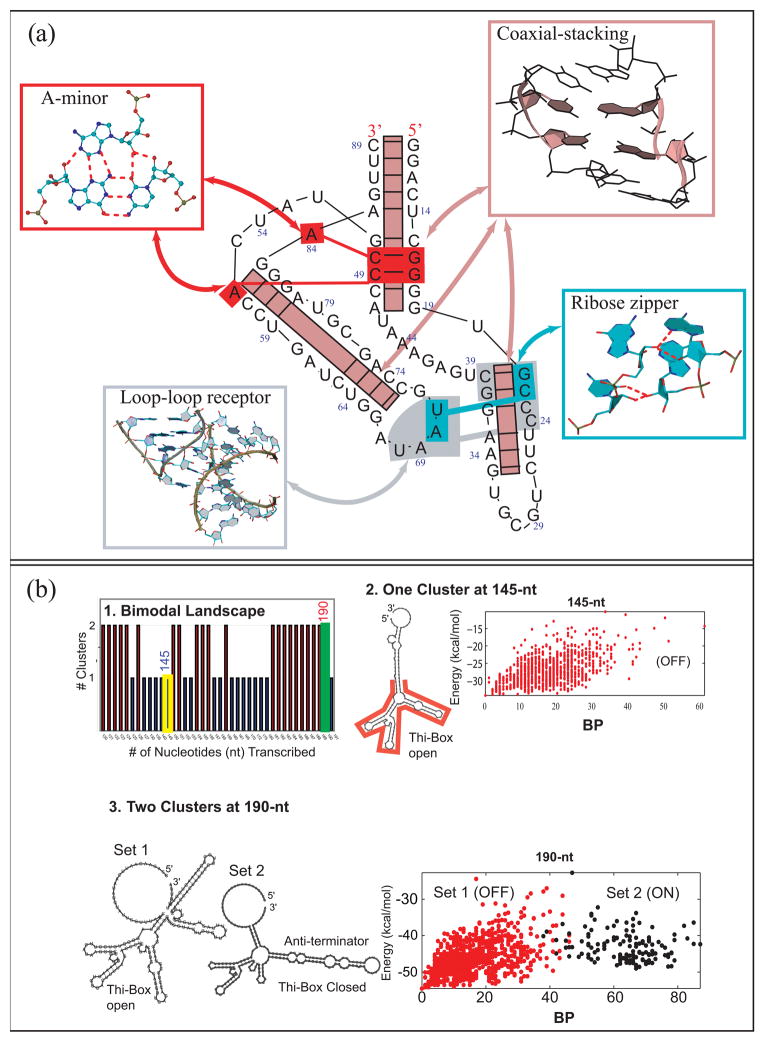
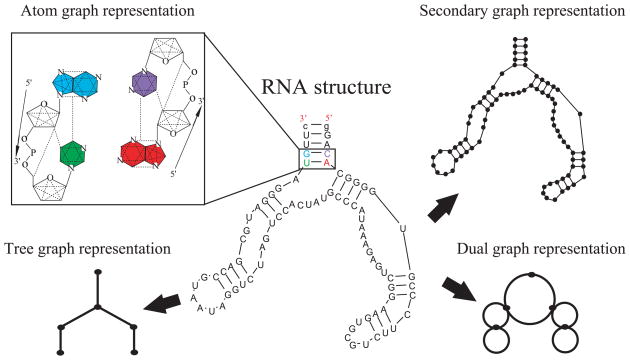
References
-
- Sioud M. Ribozymes and siRnas: from structure to preclinical applications. Handb Exp Pharmacol. 2006:223–242. - PubMed
-
- Hess H, Jaeger L. Nanobiotechnology. Curr Opin Biotechnol. 2010;21:373–375. - PubMed
-
- Laing C, Schlick T. Computational approaches to 3D modeling of RNA. Journal of Physics: Condensed Matter. 2010;22:283101. This review compares available 3D structure prediction algorithms from an RNA dataset of 43 structures of various lengths and motifs. The study finds that algorithms vary widely in terms of prediction quality; most predictions have large root-mean square deviations from the crystal structure (e.g., RMSD>6Å) due to the limitations in predicting long-range contacts. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
