

Origin and diversification dynamics of self-incompatibility haplotypes
- PMID: 21515570
- PMCID: PMC3176527
- DOI: 10.1534/genetics.111.127399
Origin and diversification dynamics of self-incompatibility haplotypes
Abstract
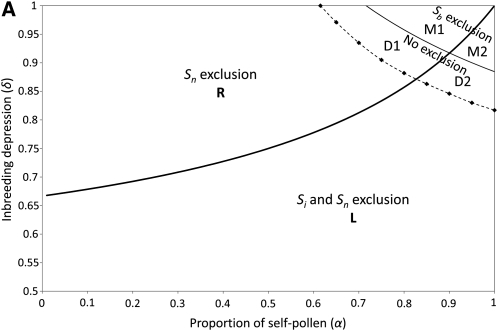
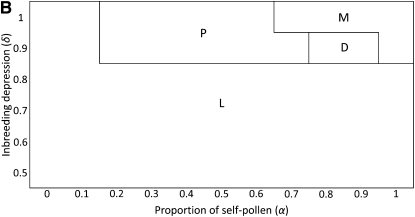
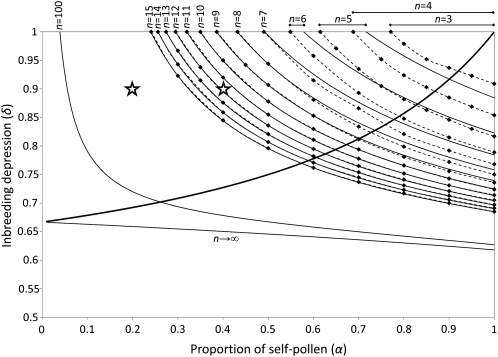
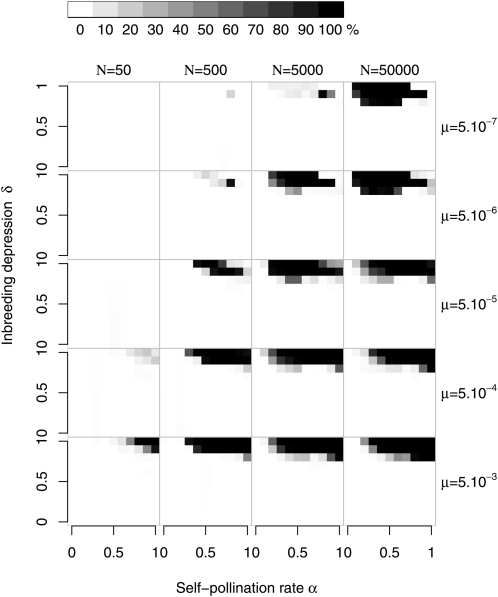
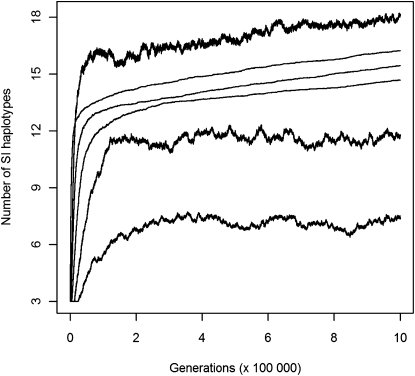
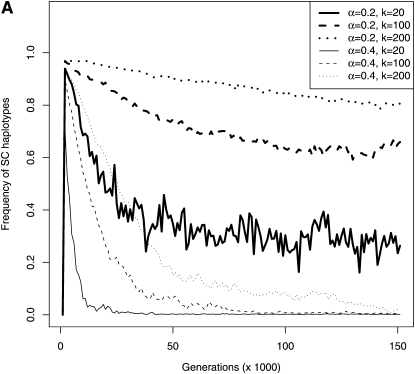
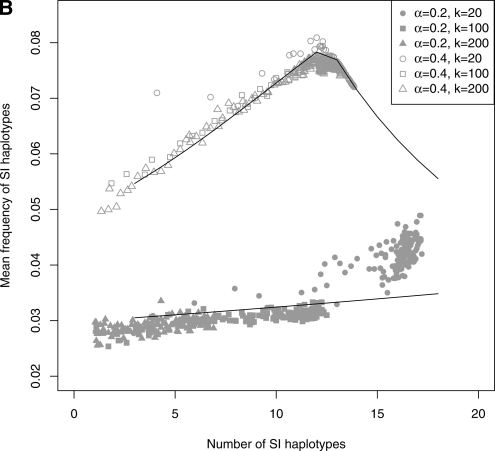
Self-incompatibility (SI) is a genetic system found in some hermaphrodite plants. Recognition of pollen by pistils expressing cognate specificities at two linked genes leads to rejection of self pollen and pollen from close relatives, i.e., to avoidance of self-fertilization and inbred matings, and thus increased outcrossing. These genes generally have many alleles, yet the conditions allowing the evolution of new alleles remain mysterious. Evolutionary changes are clearly necessary in both genes, since any mutation affecting only one of them would result in a nonfunctional self-compatible haplotype. Here, we study diversification at the S-locus (i.e., a stable increase in the total number of SI haplotypes in the population, through the incorporation of new SI haplotypes), both deterministically (by investigating analytically the fate of mutations in an infinite population) and by simulations of finite populations. We show that the conditions allowing diversification are far less stringent in finite populations with recurrent mutations of the pollen and pistil genes, suggesting that diversification is possible in a panmictic population. We find that new SI haplotypes emerge fastest in populations with few SI haplotypes, and we discuss some implications for empirical data on S-alleles. However, allele numbers in our simulations never reach values as high as observed in plants whose SI systems have been studied, and we suggest extensions of our models that may reconcile the theory and data.
Figures
Similar articles
-
How Have Self-Incompatibility Haplotypes Diversified? Generation of New Haplotypes during the Evolution of Self-Incompatibility from Self-Compatibility.Am Nat. 2016 Aug;188(2):163-74. doi: 10.1086/687110. Epub 2016 Jun 2. Am Nat. 2016. PMID: 27420782
-
Initial invasion of gametophytic self-incompatibility alleles in the absence of tight linkage between pollen and pistil S alleles.Am Nat. 2014 Aug;184(2):248-57. doi: 10.1086/676942. Epub 2014 Jul 1. Am Nat. 2014. PMID: 25058284
-
Evolutionary Pathways for the Generation of New Self-Incompatibility Haplotypes in a Nonself-Recognition System.Genetics. 2018 Jul;209(3):861-883. doi: 10.1534/genetics.118.300748. Epub 2018 Apr 30. Genetics. 2018. PMID: 29716955 Free PMC article.
-
Plant self-incompatibility in natural populations: a critical assessment of recent theoretical and empirical advances.Mol Ecol. 2004 Oct;13(10):2873-89. doi: 10.1111/j.1365-294X.2004.02267.x. Mol Ecol. 2004. PMID: 15367105 Review.
-
Molecular mechanisms of self-incompatibility in Brassicaceae and Solanaceae.Proc Jpn Acad Ser B Phys Biol Sci. 2024;100(4):264-280. doi: 10.2183/pjab.100.014. Proc Jpn Acad Ser B Phys Biol Sci. 2024. PMID: 38599847 Free PMC article. Review.
Cited by
-
Global allele polymorphism indicates a high rate of allele genesis at a locus under balancing selection.Heredity (Edinb). 2021 Jan;126(1):163-177. doi: 10.1038/s41437-020-00358-w. Epub 2020 Aug 27. Heredity (Edinb). 2021. PMID: 32855546 Free PMC article.
-
Patterns of Polymorphism at the Self-Incompatibility Locus in 1,083 Arabidopsis thaliana Genomes.Mol Biol Evol. 2017 Aug 1;34(8):1878-1889. doi: 10.1093/molbev/msx122. Mol Biol Evol. 2017. PMID: 28379456 Free PMC article.
-
The role of promiscuous molecular recognition in the evolution of RNase-based self-incompatibility in plants.Nat Commun. 2024 Jun 7;15(1):4864. doi: 10.1038/s41467-024-49163-7. Nat Commun. 2024. PMID: 38849350 Free PMC article.
-
Helitron-like transposons contributed to the mating system transition from out-crossing to self-fertilizing in polyploid Brassica napus L.Sci Rep. 2016 Sep 21;6:33785. doi: 10.1038/srep33785. Sci Rep. 2016. PMID: 27650318 Free PMC article.
-
Overcoming self-incompatibility in grasses: a pathway to hybrid breeding.Theor Appl Genet. 2016 Oct;129(10):1815-29. doi: 10.1007/s00122-016-2775-2. Epub 2016 Aug 30. Theor Appl Genet. 2016. PMID: 27577253 Review.
References
-
- Billiard S., Lopez-Villavicencio M., Devier B., Hood M., Fairhead C., Giraud T., 2011. Having sex, yes, but with whom? Inferences from fungi on the evolution of anisogamy and mating types. Biol. Rev. 86: 421–442 - PubMed
-
- Castric V., Bechsgaard J. S., Grenier S., Noureddine R., Schierup M. H., et al. , 2010. Molecular evolution within and between self-incompatibility specificities. Mol. Biol. Evol. 27: 11–20 - PubMed
-
- Charlesworth D., Charlesworth B., 1979. Evolution and breakdown of S-allele systems. Heredity 43: 41–55
MeSH terms
LinkOut - more resources
Full Text Sources
