

In vitro epithelial-to-mesenchymal transformation in human adult epicardial cells is regulated by TGFβ-signaling and WT1
- PMID: 21516490
- PMCID: PMC3149675
- DOI: 10.1007/s00395-011-0181-0
In vitro epithelial-to-mesenchymal transformation in human adult epicardial cells is regulated by TGFβ-signaling and WT1
Abstract
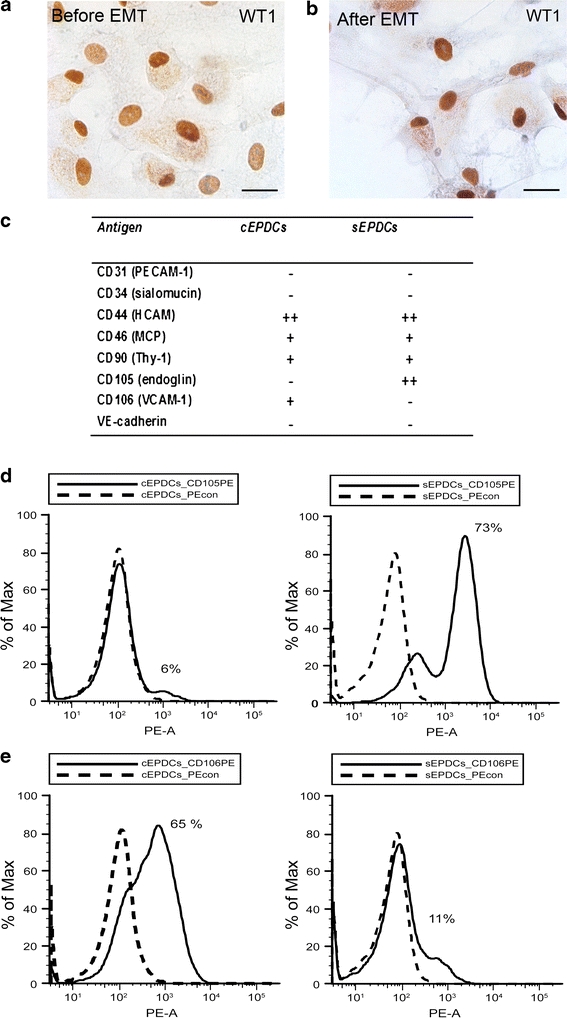
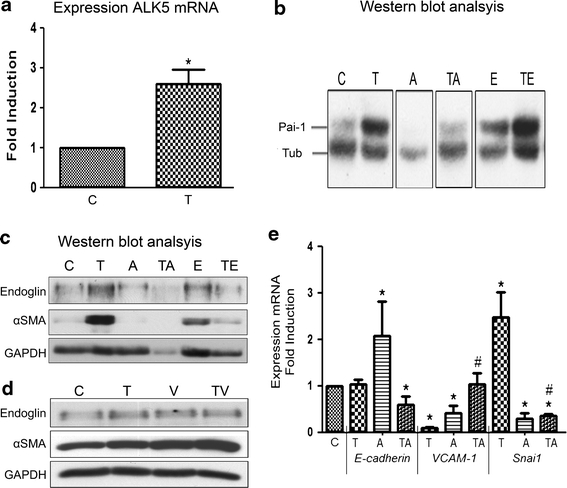
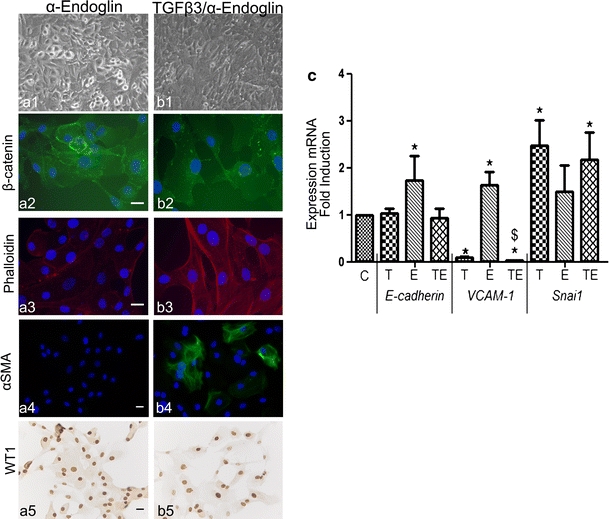
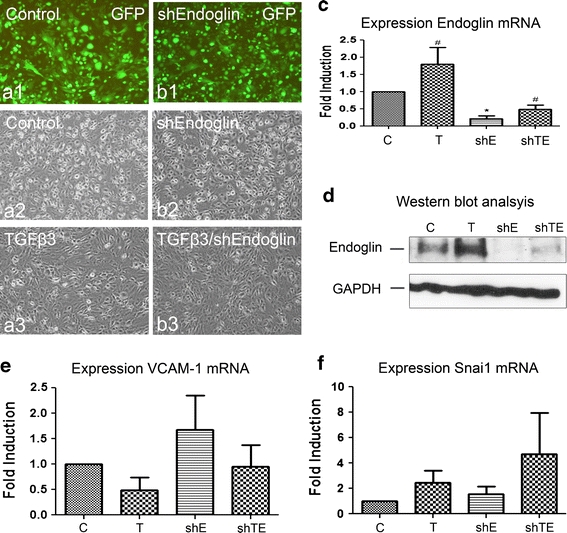
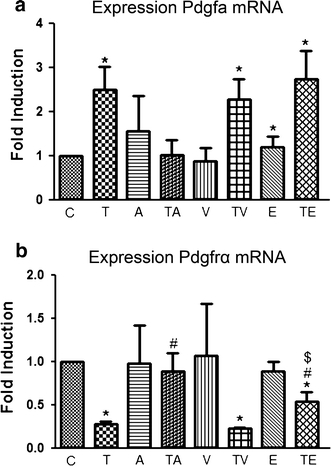
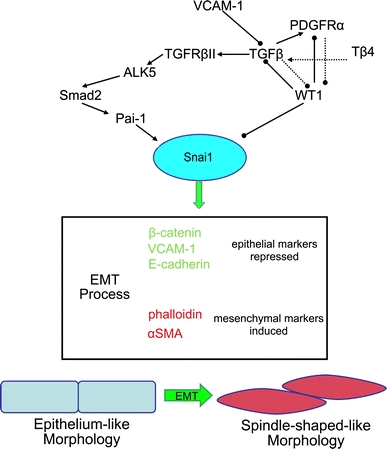
Adult epicardial cells are required for endogenous cardiac repair. After myocardial injury, they are reactivated, undergo epithelial-to-mesenchymal transformation (EMT) and migrate into the injured myocardium where they generate various cell types, including coronary smooth muscle cells and cardiac interstitial fibroblasts, which contribute to cardiac repair. To understand what drives epicardial EMT, we used an in vitro model for human adult epicardial cells. These cells have an epithelium-like morphology and markedly express the cell surface marker vascular cell adhesion marker (VCAM-1). In culture, epicardial cells spontaneously undergo EMT after which the spindle-shaped cells now express endoglin. Both epicardial cells before and after EMT express the epicardial marker, Wilms tumor 1 (WT1). Adding transforming growth factor beta (TGFβ) induces loss of epithelial character and initiates the onset of mesenchymal differentiation in human adult epicardial cells. In this study, we show that TGFβ-induced EMT is dependent on type-1 TGFβ receptor activity and can be inhibited by soluble VCAM-1. We also show that epicardial-specific knockdown of Wilms tumor-1 (WT1) induces the process of EMT in human adult epicardial cells, through transcriptional regulation of platelet-derived growth factor receptor alpha (Pdgfrα), Snai1 and VCAM-1. These data provide new insights into the process of EMT in human adult epicardial cells, which might provide opportunities to develop new strategies for endogenous cell-based cardiac repair.
Figures





References
-
- Al-Nedawi KN, Czyz M, Bednarek R, Szemraj J, Swiatkowska M, Cierniewska-Cieslak A, Wyczolkowska J, Cierniewski CS. Thymosin beta 4 induces the synthesis of plasminogen activator inhibitor 1 in cultured endothelial cells and increases its extracellular expression. Blood. 2004;103:1319–1324. doi: 10.1182/blood-2003-04-1015. - DOI - PubMed
-
- Arthur HM, Ure J, Smith AJ, Renforth G, Wilson DI, Torsney E, Charlton R, Parums DV, Jowett T, Marchuk DA, Burn J, Diamond AG. Endoglin, an ancillary TGFbeta receptor, is required for extraembryonic angiogenesis and plays a key role in heart development. Dev Biol. 2000;217:42–53. doi: 10.1006/dbio.1999.9534. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
