

Tapasin discriminates peptide-human leukocyte antigen-A*02:01 complexes formed with natural ligands
- PMID: 21518758
- PMCID: PMC3121515
- DOI: 10.1074/jbc.M111.230151
Tapasin discriminates peptide-human leukocyte antigen-A*02:01 complexes formed with natural ligands
Abstract
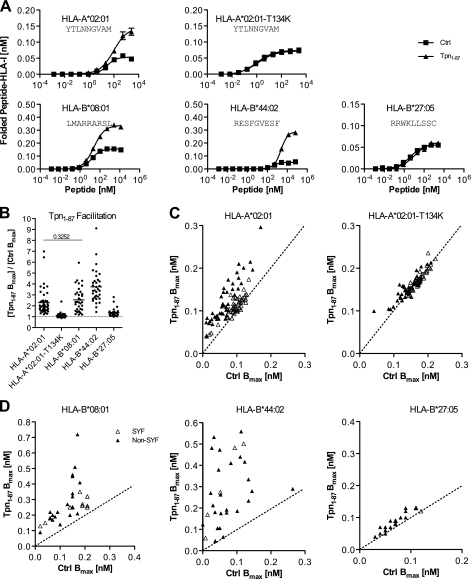
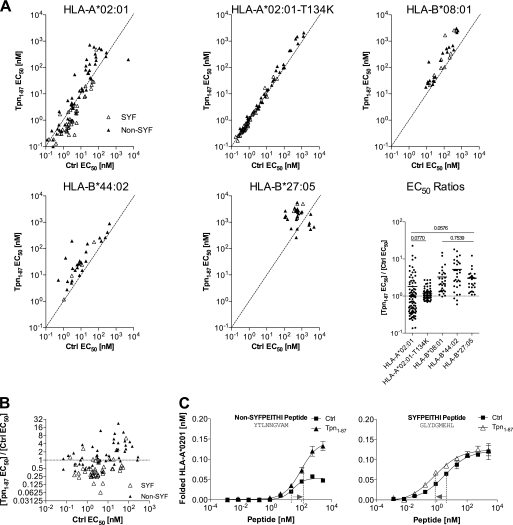
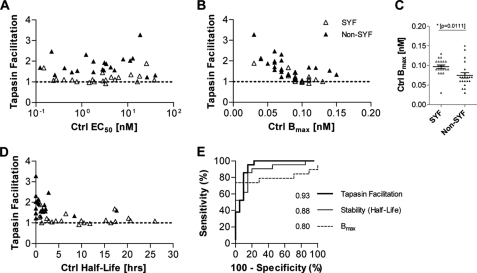
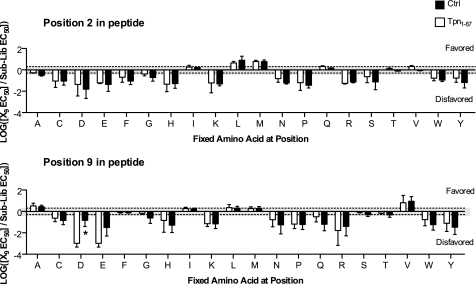
A plethora of peptides are generated intracellularly, and most peptide-human leukocyte antigen (HLA)-I interactions are of a transient, unproductive nature. Without a quality control mechanism, the HLA-I system would be stressed by futile attempts to present peptides not sufficient for the stable peptide-HLA-I complex formation required for long term presentation. Tapasin is thought to be central to this essential quality control, but the underlying mechanisms remain unknown. Here, we report that the N-terminal region of tapasin, Tpn(1-87), assisted folding of peptide-HLA-A*02:01 complexes according to the identity of the peptide. The facilitation was also specific for the identity of the HLA-I heavy chain, where it correlated to established tapasin dependence hierarchies. Two large sets of HLA-A*02:01 binding peptides, one extracted from natural HLA-I ligands from the SYFPEITHI database and one consisting of medium to high affinity non-SYFPEITHI ligands, were studied in the context of HLA-A*02:01 binding and stability. We show that the SYFPEITHI peptides induced more stable HLA-A*02:01 molecules than the other ligands, although affinities were similar. Remarkably, Tpn(1-87) could functionally discriminate the selected SYFPEITHI peptides from the other peptide binders with high sensitivity and specificity. We suggest that this HLA-I- and peptide-specific function, together with the functions exerted by the more C-terminal parts of tapasin, are major features of tapasin-mediated HLA-I quality control. These findings are important for understanding the biogenesis of HLA-I molecules, the selection of presented T-cell epitopes, and the identification of immunogenic targets in both basic research and vaccine design.
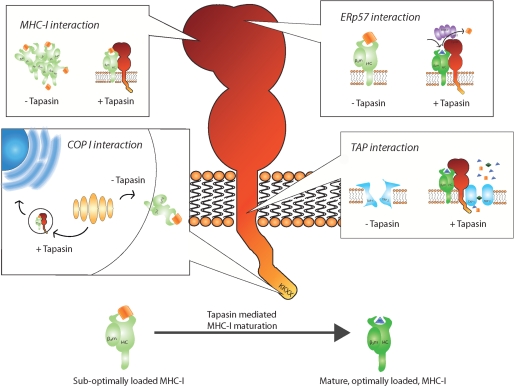
Figures
References
-
- Cresswell P., Bangia N., Dick T., Diedrich G. (1999) Immunol. Rev. 172, 21–28 - PubMed
-
- Park B., Lee S., Kim E., Cho K., Riddell S. R., Cho S., Ahn K. (2006) Cell 127, 369–382 - PubMed
-
- Ortmann B., Copeman J., Lehner P. J., Sadasivan B., Herberg J. A., Grandea A. G., Riddell S. R., Tampé R., Spies T., Trowsdale J., Cresswell P. (1997) Science 277, 1306–1309 - PubMed
-
- Grandea A. G., 3rd, Lehner P. J., Cresswell P., Spies T. (1997) Immunogenetics 46, 477–483 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
