

Estradiol and progesterone strongly inhibit the innate immune response of mononuclear cells in newborns
- PMID: 21518785
- PMCID: PMC3191988
- DOI: 10.1128/IAI.00076-11
Estradiol and progesterone strongly inhibit the innate immune response of mononuclear cells in newborns
Abstract
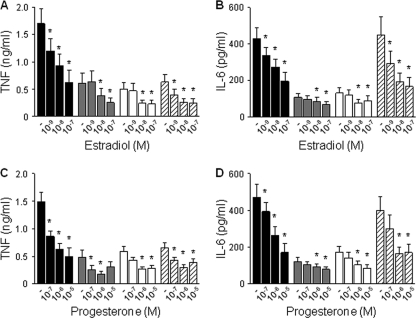
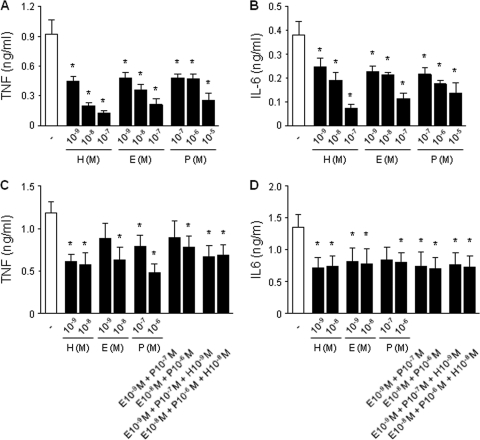
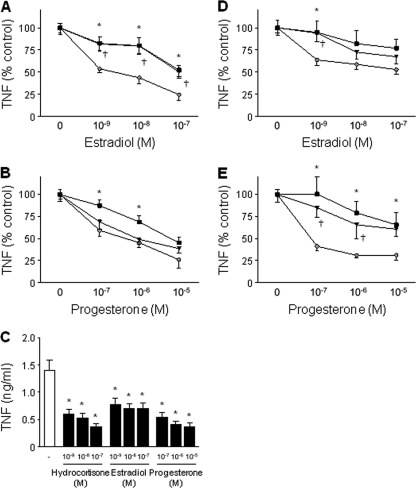
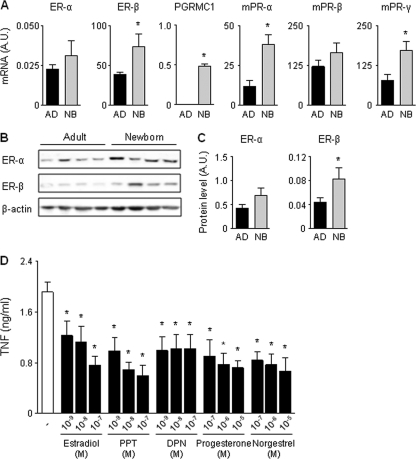
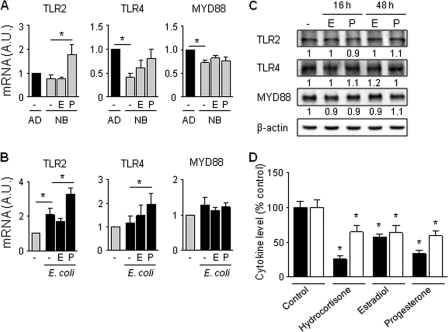
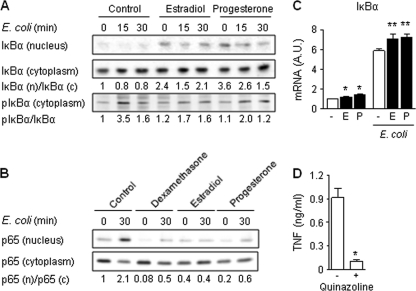
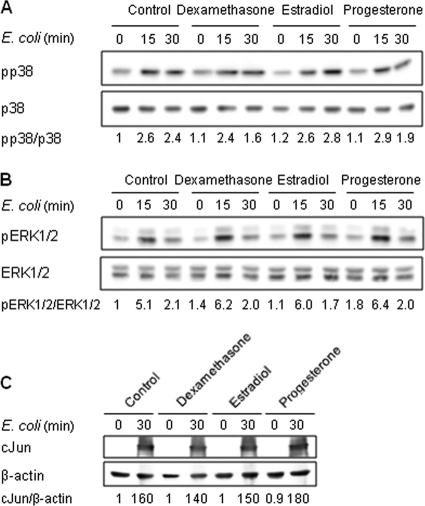
Newborns are particularly susceptible to bacterial infections due to qualitative and quantitative deficiencies of the neonatal innate immune system. However, the mechanisms underlying these deficiencies are poorly understood. Given that fetuses are exposed to high concentrations of estradiol and progesterone during gestation and at time of delivery, we analyzed the effects of these hormones on the response of neonatal innate immune cells to endotoxin, bacterial lipopeptide, and Escherichia coli and group B Streptococcus, the two most common causes of early-onset neonatal sepsis. Here we show that at concentrations present in umbilical cord blood, estradiol and progesterone are as powerful as hydrocortisone for inhibition of cytokine production by cord blood mononuclear cells (CBMCs) and newborn monocytes. Interestingly, CBMCs and newborn monocytes are more sensitive to the effects of estradiol and progesterone than adult peripheral blood mononuclear cells and monocytes. This increased sensitivity is associated with higher expression levels of estrogen and membrane progesterone receptors but is independent of a downregulation of Toll-like receptor 2 (TLR2), TLR4, and myeloid differentiation primary response gene 88 in newborn cells. Estradiol and progesterone mediate their anti-inflammatory activity through inhibition of the NF-κB pathway but not the mitogen-activated protein kinase pathway in CBMCs. Altogether, these results suggest that elevated umbilical cord blood concentrations of estradiol and progesterone acting on mononuclear cells expressing high levels of steroid receptors contribute to impair innate immune responses in newborns. Therefore, intrauterine exposure to estradiol and progesterone may participate in increasing susceptibility to infection during the neonatal period.
Figures
Similar articles
-
Female sex hormones modulate Porphyromonas gingivalis lipopolysaccharide-induced Toll-like receptor signaling in primary human monocytes.J Periodontal Res. 2016 Jun;51(3):395-406. doi: 10.1111/jre.12320. Epub 2015 Sep 14. J Periodontal Res. 2016. PMID: 26364725
-
Human neonatal peripheral blood leukocytes demonstrate pathogen-specific coordinate expression of TLR2, TLR4/MD2, and MyD88 during bacterial infection in vivo.Pediatr Res. 2010 Dec;68(6):479-83. doi: 10.1203/PDR.0b013e3181f90810. Pediatr Res. 2010. PMID: 20805788 Free PMC article.
-
Pellino-3 promotes endotoxin tolerance and acts as a negative regulator of TLR2 and TLR4 signaling.J Leukoc Biol. 2015 Dec;98(6):963-74. doi: 10.1189/jlb.2VMA0515-229RR. Epub 2015 Aug 26. J Leukoc Biol. 2015. PMID: 26310831 Free PMC article.
-
An impaired inflammatory cytokine response to gram-negative LPS in human neonates is associated with the defective TLR-mediated signaling pathway.J Clin Immunol. 2015 Feb;35(2):218-26. doi: 10.1007/s10875-015-0128-6. Epub 2015 Jan 29. J Clin Immunol. 2015. PMID: 25631527
-
Increased toll-like receptor (TLR) 2 and TLR4 expression in monocytes from patients with type 1 diabetes: further evidence of a proinflammatory state.J Clin Endocrinol Metab. 2008 Feb;93(2):578-83. doi: 10.1210/jc.2007-2185. Epub 2007 Nov 20. J Clin Endocrinol Metab. 2008. PMID: 18029454 Free PMC article.
Cited by
-
Human monocyte subtype expression of neuroinflammation- and regeneration-related genes is linked to age and sex.PLoS One. 2024 Oct 30;19(10):e0300946. doi: 10.1371/journal.pone.0300946. eCollection 2024. PLoS One. 2024. PMID: 39475881 Free PMC article.
-
Multidrug resistance crisis during COVID-19 pandemic: Role of anti-microbial peptides as next-generation therapeutics.Colloids Surf B Biointerfaces. 2022 Mar;211:112303. doi: 10.1016/j.colsurfb.2021.112303. Epub 2021 Dec 20. Colloids Surf B Biointerfaces. 2022. PMID: 34952285 Free PMC article. Review.
-
High expression levels of macrophage migration inhibitory factor sustain the innate immune responses of neonates.Proc Natl Acad Sci U S A. 2016 Feb 23;113(8):E997-1005. doi: 10.1073/pnas.1514018113. Epub 2016 Feb 8. Proc Natl Acad Sci U S A. 2016. PMID: 26858459 Free PMC article.
-
Sex-related immunity: could Toll-like receptors be the answer in acute inflammatory response?Front Immunol. 2024 May 21;15:1379754. doi: 10.3389/fimmu.2024.1379754. eCollection 2024. Front Immunol. 2024. PMID: 38835761 Free PMC article. Review.
-
Resolution of Crohn's disease and complex regional pain syndrome following treatment of paratuberculosis.World J Gastroenterol. 2015 Apr 7;21(13):4048-62. doi: 10.3748/wjg.v21.i13.4048. World J Gastroenterol. 2015. PMID: 25852293 Free PMC article.
References
-
- Adkins B., Leclerc C., Marshall-Clarke S. 2004. Neonatal adaptive immunity comes of age. Nat. Rev. Immunol. 4:553–564 - PubMed
-
- Asai K., et al. 2001. Gender differences in cytokine secretion by human peripheral blood mononuclear cells: role of estrogen in modulating LPS-induced cytokine secretion in an ex vivo septic model. Shock 16:340–343 - PubMed
-
- Bessler H., et al. 1999. Effects of dexamethasone on IL-1beta, IL-6, and TNF-alpha production by mononuclear cells of newborns and adults. Biol. Neonate 75:225–233 - PubMed
-
- Bromberger P., et al. 2000. The influence of intrapartum antibiotics on the clinical spectrum of early-onset group B streptococcal infection in term infants. Pediatrics 106:244–250 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
