

A barrier-only boundary element delimits the formation of facultative heterochromatin in Drosophila melanogaster and vertebrates
- PMID: 21518956
- PMCID: PMC3133385
- DOI: 10.1128/MCB.05165-11
A barrier-only boundary element delimits the formation of facultative heterochromatin in Drosophila melanogaster and vertebrates
Abstract
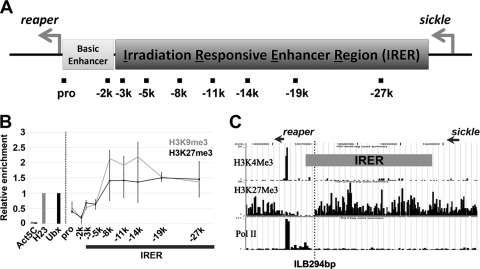
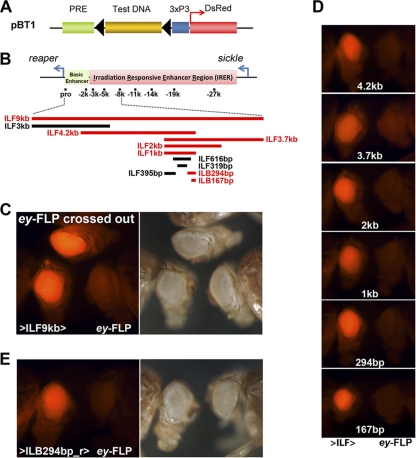
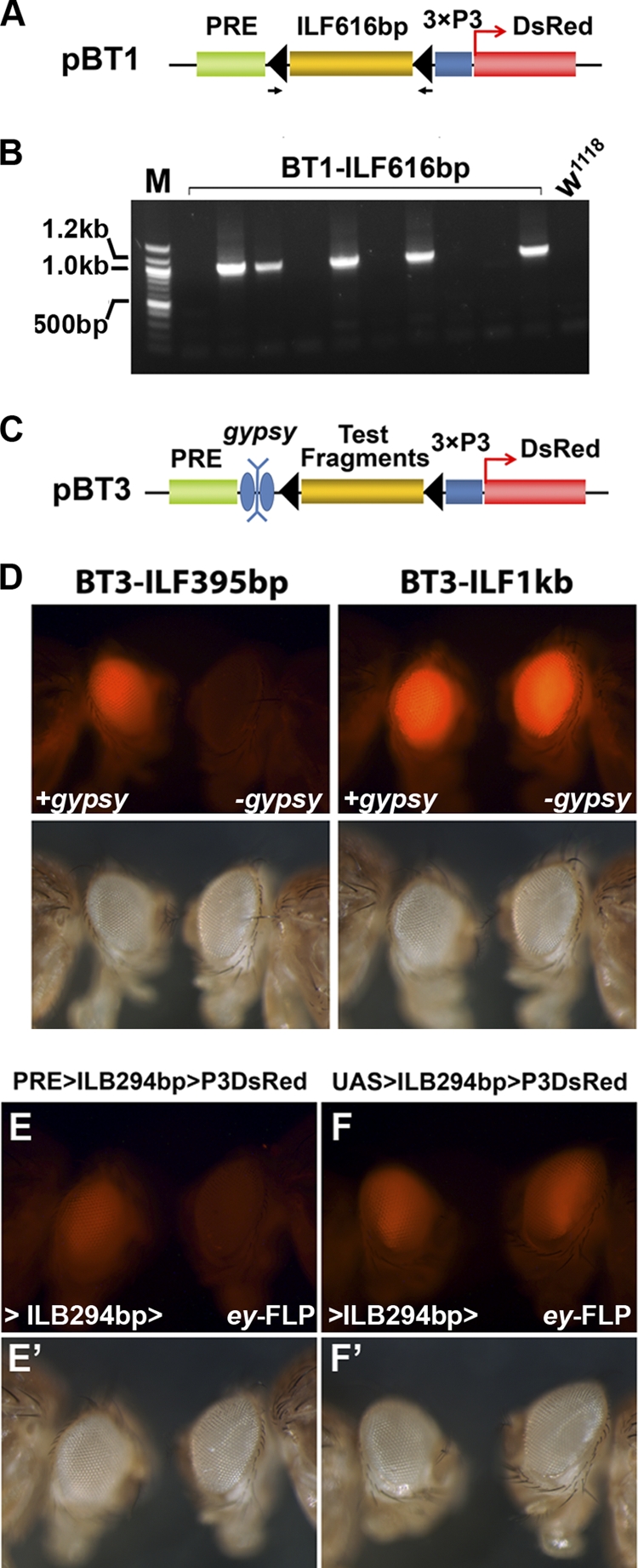
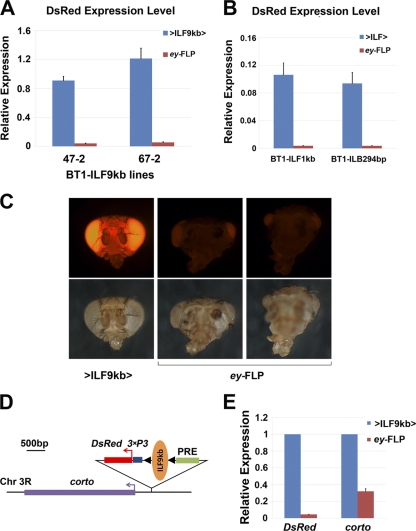
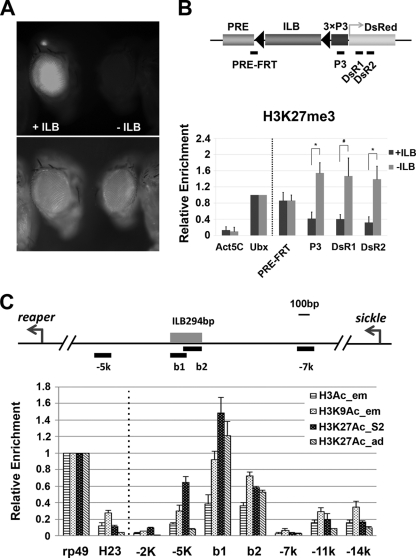
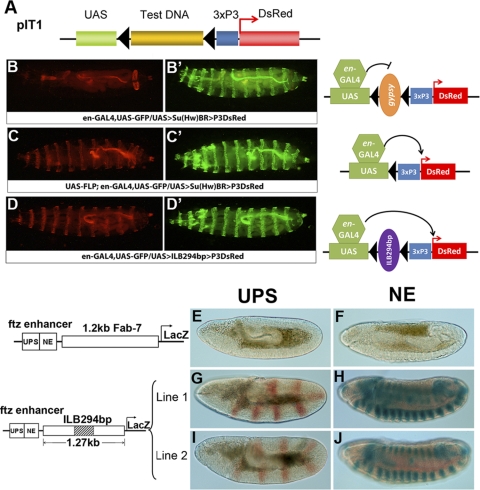
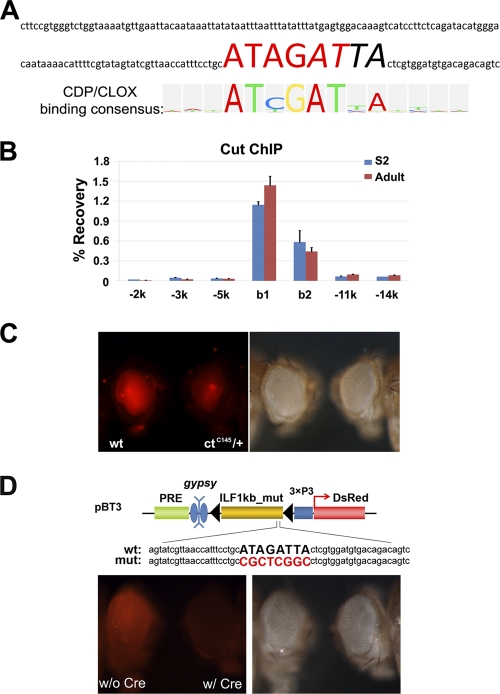
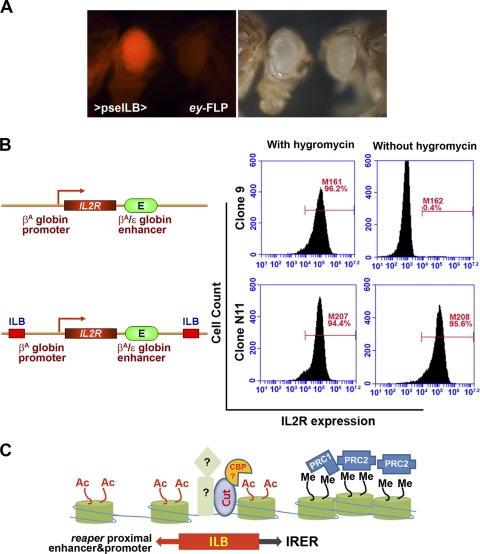
Formation of facultative heterochromatin at specific genomic loci is fundamentally important in defining cellular properties such as differentiation potential and responsiveness to developmental, physiological, and environmental stimuli. By the nature of their formation, heterochromatin and repressive histone marks propagate until the chain reaction is broken. While certain active promoters can block propagation of heterochromatin, there are also specialized DNA elements, referred to as chromatin barriers, that serve to demarcate the boundary of facultative heterochromatin formation. In this study, we identified a chromatin barrier that specifically limits the formation of repressive chromatin to a distal enhancer region so that repressive histone modifications cannot reach the promoter and promoter-proximal enhancer regions of reaper. Unlike all of the known boundary elements identified for Drosophila melanogaster, this IRER (irradiation-responsive enhancer region) left barrier (ILB) does not exhibit enhancer-blocking activity. Not only has the ILB been conserved in different Drosophila species, it can also function as an effective chromatin barrier in vertebrate cells. This suggests that the mechanism by which it functions to spatially restrict the formation of repressive chromatin marked by trimethylated H3K27 has also been conserved widely during evolution.
Figures
References
-
- Andres V., Chiara M. D., Mahdavi V. 1994. A new bipartite DNA-binding domain: cooperative interaction between the cut repeat and homeo domain of the cut homeo proteins. Genes Dev. 8:245–257 - PubMed
-
- Bell A. C., West A. G., Felsenfeld G. 1999. The protein CTCF is required for the enhancer blocking activity of vertebrate insulators. Cell 98:387–396 - PubMed
-
- Bello B. C., Hirth F., Gould A. P. 2003. A pulse of the Drosophila Hox protein Abdominal-A schedules the end of neural proliferation via neuroblast apoptosis. Neuron 37:209–219 - PubMed
-
- Bi X., Broach J. R. 2001. Chromosomal boundaries in S. cerevisiae. Curr. Opin. Genet. Dev. 11:199–204 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases