

A mutation in the dynein heavy chain gene compensates for energy deficit of mutant SOD1 mice and increases potentially neuroprotective IGF-1
- PMID: 21521523
- PMCID: PMC3111394
- DOI: 10.1186/1750-1326-6-26
A mutation in the dynein heavy chain gene compensates for energy deficit of mutant SOD1 mice and increases potentially neuroprotective IGF-1
Abstract
Background: Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease characterized by a progressive loss of motor neurons. ALS patients, as well as animal models such as mice overexpressing mutant SOD1s, are characterized by increased energy expenditure. In mice, this hypermetabolism leads to energy deficit and precipitates motor neuron degeneration. Recent studies have shown that mutations in the gene encoding the dynein heavy chain protein are able to extend lifespan of mutant SOD1 mice. It remains unknown whether the protection offered by these dynein mutations relies on a compensation of energy metabolism defects.
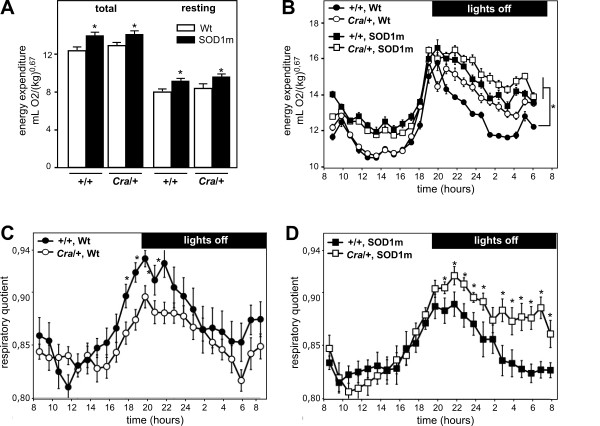
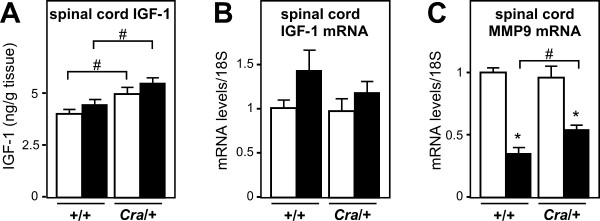
Results: SOD1(G93A) mice were crossbred with mice harboring the dynein mutant Cramping allele (Cra/+ mice). Dynein mutation increased adipose stores in compound transgenic mice through increasing carbohydrate oxidation and sparing lipids. Metabolic changes that occurred in double transgenic mice were accompanied by the normalization of the expression of key mRNAs in the white adipose tissue and liver. Furthermore, Dynein Cra mutation rescued decreased post-prandial plasma triglycerides and decreased non esterified fatty acids upon fasting. In SOD1(G93A) mice, the dynein Cra mutation led to increased expression of IGF-1 in the liver, increased systemic IGF-1 and, most importantly, to increased spinal IGF-1 levels that are potentially neuroprotective.
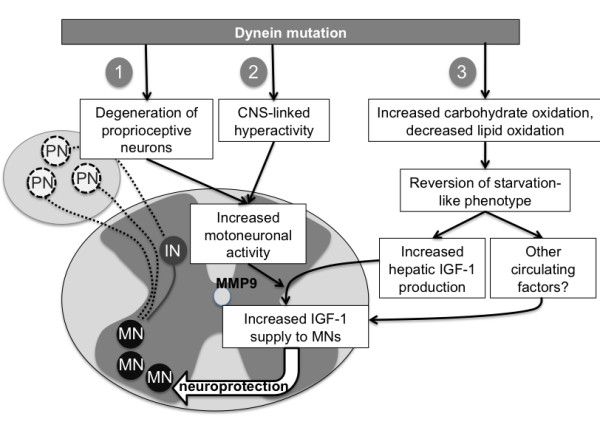
Conclusions: These findings suggest that the protection against SOD1(G93A) offered by the Cramping mutation in the dynein gene is, at least partially, mediated by a reversal in energy deficit and increased IGF-1 availability to motor neurons.
Figures



References
-
- Dupuis L, Pradat PF, Ludolph AC, Loeffler JP. Energy metabolism in amyotrophic lateral sclerosis. Lancet Neurol. 2010;10(1):75–82. - PubMed
-
- Dupuis L, Corcia P, Fergani A, Gonzalez De Aguilar JL, Bonnefont-Rousselot D, Bittar R, Seilhean D, Hauw JJ, Lacomblez L, Loeffler JP, Meininger V. Dyslipidemia is a protective factor in amyotrophic lateral sclerosis. Neurology. 2008;70:1004–1009. doi: 10.1212/01.wnl.0000285080.70324.27. - DOI - PubMed
-
- Dorst J, Kühnlein P, Hendrich C, Kassubek J, Sperfeld AD, Ludolph AC. Patients with elevated triglyceride and cholesterol serum levels have a prolonged survival in Amyotrophic Lateral Sclerosis. J Neurol. 2010;258(4):613–7. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
