

The oriented emergence of axons from retinal ganglion cells is directed by laminin contact in vivo
- PMID: 21521613
- PMCID: PMC3087191
- DOI: 10.1016/j.neuron.2011.03.013
The oriented emergence of axons from retinal ganglion cells is directed by laminin contact in vivo
Abstract
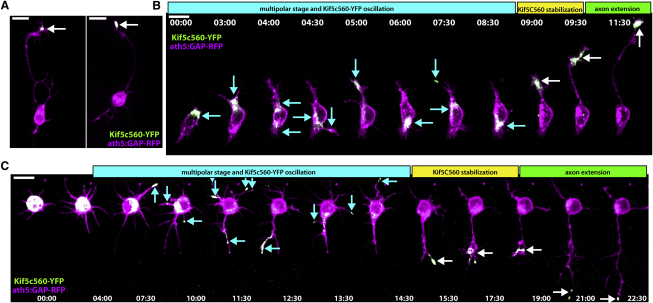
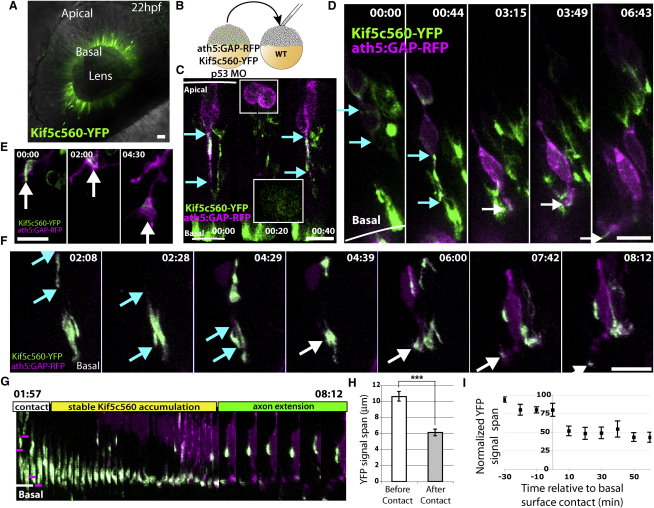
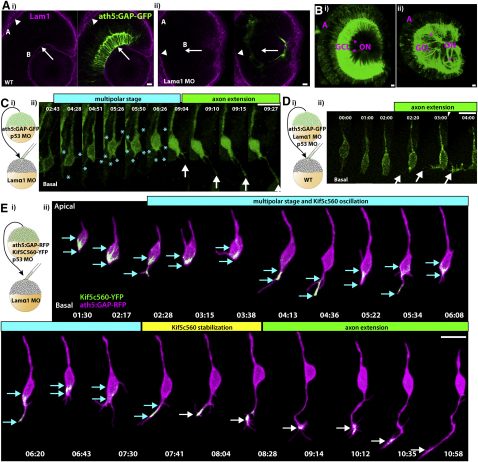
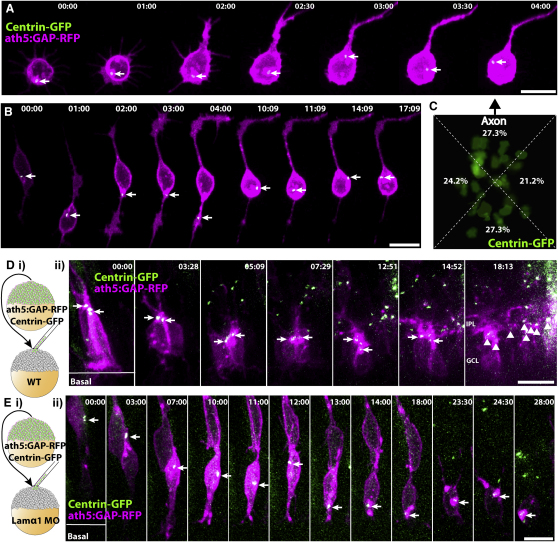
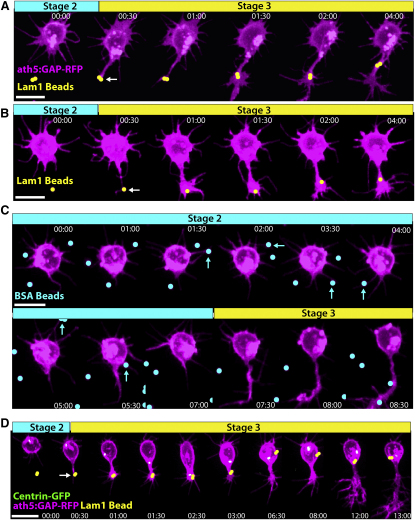
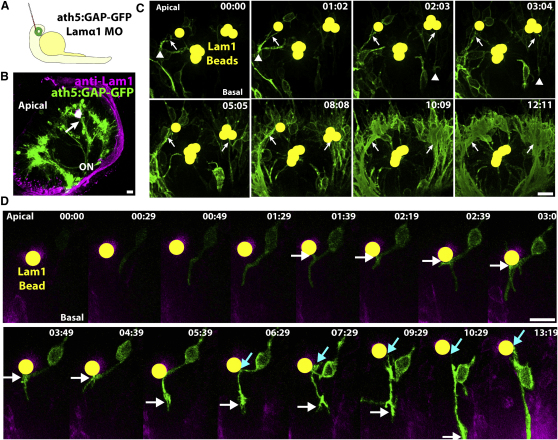
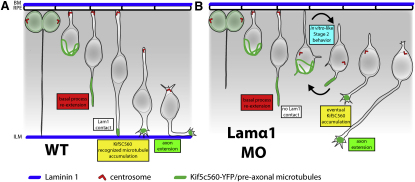
How the site of axon emergence is specified during neural development is not understood. Previous studies disagree on the relative importance of intrinsic and extrinsic mechanisms. The axons of retinal ganglion cells (RGCs) emerge basally in vivo, yet because RGCs develop from polarized neuroepithelial cells within a polarized environment, disentangling intrinsic and extrinsic influences is a challenge. We use time-lapse imaging to demonstrate that Laminin acting directly on RGCs is necessary and sufficient to orient axon emergence in vivo. Laminin contact with the basal processes of newborn RGCs prevents the cells from entering a stochastic Stage 2 phase, directs the rapid accumulation of the early axonal marker Kif5c560-YFP, and leads to the formation of axonal growth cones. These results suggest that contact-mediated cues may be critical for the site of axon emergence and account for the differences in cellular behavior observed in vitro and in vivo.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures

References
-
- Arimura N., Kaibuchi K. Neuronal polarity: from extracellular signals to intracellular mechanisms. Nat. Rev. Neurosci. 2007;8:194–205. - PubMed
-
- Barnes A.P., Lilley B.N., Pan Y.A., Plummer L.J., Powell A.W., Raines A.N., Sanes J.R., Polleux F. LKB1 and SAD kinases define a pathway required for the polarization of cortical neurons. Cell. 2007;129:549–563. - PubMed
-
- Basto R., Lau J., Vinogradova T., Gardiol A., Woods C.G., Khodjakov A., Raff J.W. Flies without centrioles. Cell. 2006;125:1375–1386. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
