

Review
doi: 10.1242/dev.054080.
Principles of planar polarity in animal development
Affiliations
- PMID: 21521735
- PMCID: PMC3082295
- DOI: 10.1242/dev.054080
Item in Clipboard
Review
Principles of planar polarity in animal development
Development.
2011 May.
Abstract
Planar polarity describes the coordinated polarisation of cells or structures in the plane of a tissue. The patterning mechanisms that underlie planar polarity are well characterised in Drosophila, where many events are regulated by two pathways: the 'core' planar polarity complex and the Fat/Dachsous system. Components of both pathways also function in vertebrates and are implicated in diverse morphogenetic processes, some of which self-evidently involve planar polarisation and some of which do not. Here, we review the molecular mechanisms and cellular consequences of planar polarisation in diverse contexts, seeking to identify the common principles across the animal kingdom.
Figures
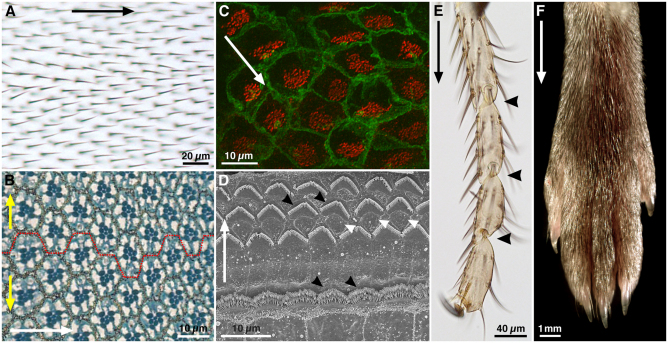
Examples of planar polarity in flies and mice. (A) Adult Drosophila wing surface. Planar polarity is evident in the organization of trichomes on the proximodistal (PD) axis (black arrow). (B) Sub-apical section through an adult Drosophila eye at the dorsoventral (DV) midline. Each eye facet comprises a group of ~20 cells (an ommatidium). In this section plane, the pigmented rhabdomeres (dark blue) of seven photoreceptors are in the centre of each ommatidium. Ommatidia are hexagonally tesselated and show mirror-image symmetry around the DV midline (broken red line), revealing axes of planar polarity on the anteroposterior (AP, white arrow) and DV (yellow arrows) axes. (C) Confocal image of mouse brain ependymal cells, with cell membranes stained for β-catenin (green) and cilia basal bodies for γ-tubulin (red). In each cell, basal bodies are displaced towards one side (white arrow), creating ‘translational’ polarity. (D) Scanning electron micrograph of an adult mouse organ of Corti. Planar polarity is seen on the mediolateral axis (white arrow), in the arrangement of the hair cell stereocilia (black arrowheads). Support cells between the hair cells (white arrowheads) show no overt morphological polarisation. (E) Tarsal joints of an adult Drosophila leg. Planar polarity is evident on the PD axis (black arrow) through the polarisation of bristles and the joints (black arrowheads). (F) Distal end of an adult mouse leg, showing the PD polarised arrangement of the fur. Images courtesy of Dr Henry Ho (C), Dr XuDong Wu (D) and Dr Cindy Lu (F), unpublished. (A,B,E) D.S., unpublished.
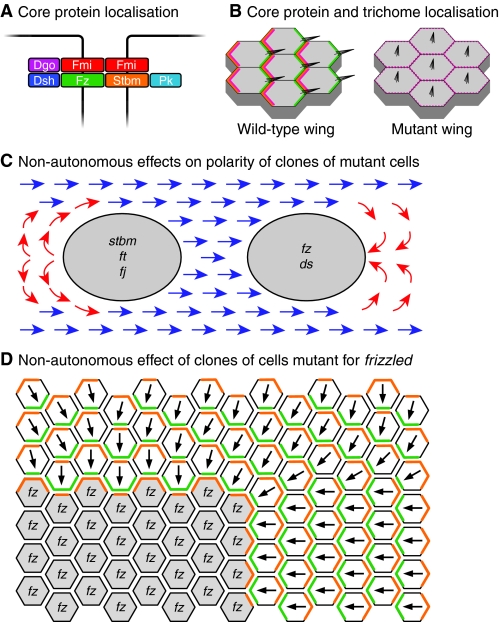
Properties of the core planar polarity proteins in Drosophila wing development. (A) Core protein arrangement at the adherens junction zone of epithelial cells in the Drosophila wing. An intercellular asymmetric junctional complex forms, with the transmembrane proteins Fz (green) and Fmi (red), and the cytosolic proteins Dsh (dark blue) and Dgo (purple) in one cell, associating with the transmembrane proteins Stbm (orange) and Fmi, and the cytosolic protein Pk (pale blue) in the adjacent cell. (B) Subcellular distribution of the core proteins and effectors in the pupal wing. From ~28 hours of pupal life, the core proteins show strong asymmetric subcellular distributions, with Fz, Dsh, Dgo and Fmi (green) being localised at distal cell edges (right), and Stbm, Pk and Fmi (orange) at proximal cell edges (left). The effectors Inturned, Fuzzy, Fritz and Multiple Wing Hairs are recruited proximally (pink), and locally inhibit trichome formation, such that trichomes (black) only emerge distally. Cells that lack or have uniform core protein activity show unpolarised effector protein localisation (pink) and trichome production in the cell centre. (C) Non-autonomous effects on trichome polarity (normal polarity shown in blue) in the wing caused by clones of cells lacking planar polarity gene function. Groups of cells lacking stbm, ft or fj activity (grey, left) cause trichomes proximal to the clone to invert their polarity (red arrows), whereas groups of cells lacking fz or ds function (grey, right) cause trichomes distal to the clone to invert their polarity. (D) Model of how a fz clone might alter trichome polarity in neighbouring wild-type cells. Cells lacking Fz (grey) can only form complexes containing Stbm (orange) inside the clone, interacting with Fz (green) in the neighbouring wild-type cells. The abnormal polarity of these junctional complexes around the edge of the clone then propagates to neighbouring cells. Dgo, Diego; ds, dachsous; Dsh, Dishevelled; fj, four-jointed; Fmi, Flamingo; ft, fat; Fz, Frizzled; Pk, Prickle; Stbm, Strabismus.

Fat and Dachsous interactions in the Drosophila wing. (A,B) Schematics of the interactions between Fat (Ft) and Dachsous (Ds) at adherens junctions of epithelial cells in the Drosophila imaginal discs. (A) Ft (cyan) and Ds (magenta) are large atypical cadherin molecules that interact heterophilically. (B) Ft/Ds heterophilic interactions are modulated by Four-jointed (Fj, yellow) activity in the Golgi, which phosphorylates the extracellular cadherin repeats in Ft and Ds as they traffic to the cell surface. Fj-mediated phosphorylation of Ft increases its binding affinity for Ds, whereas phosphorylation of Ds decreases its affinity for Ft.
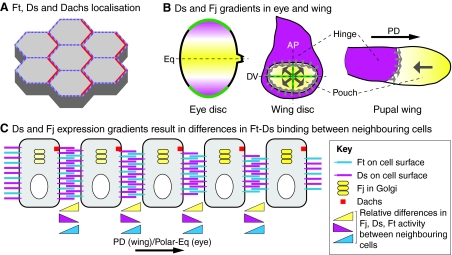
Properties of the Fat and Dachsous system in Drosophila. (A) A schematic of the apical surface of epithelial cells in the Drosophila wing. Fat (Ft, blue) and Dachsous (Ds, magenta) are uniformly distributed, but promote planar polarised asymmetry of Dachs (red). (B) Ds and Four-jointed (Fj) gradients. In the eye disc (left), the secreted morphogen Wg (green) is highly expressed at the dorsal and ventral poles, and activates Ds (magenta) expression while suppressing Fj (yellow) expression. The dotted line indicates dorsoventral (DV) midline, also known as the equator (Eq). In the third instar wing disc (middle) and early pupal wing (right), high levels of Ds are present outside the wing pouch, including the hinge. Fj shows graded expression (large grey arrows) within the wing pouch that is highest distally, probably owing to morphogen signalling from the DV and anteroposterior (AP) midlines (green). The sharp boundary between high Ds hinge expression and low Fj pouch expression might serve as a polarising cue (small grey arrows). (C) Model of how opposing gradients of Ds and Fj expression could lead to different levels of Ft/Ds binding activity across cells on the proximodistal (PD) axis of the wing disc or the polar-equatorial (Polar-Eq) axis of the eye disc. Ds levels decline left to right, Fj levels decline right to left and Ft levels are uniform. Triangles at the bottom indicate relative differences between Ds, Ft and Fj activity between pairs of cells. Fj cell-autonomously inhibits Ds activity and simultaneously enhances Ft activity. Thus, considering any cell, its left neighbour has higher Ds expression and activity and lower Ft activity than its right neighbour, promoting formation of more and stronger heterophilic Ft-Ds interactions on the left-right axis and fewer and weaker interactions on the right-left axis. The resulting difference in the strengths of Ft-Ds interactions on opposing cell edges leads to asymmetric distribution of Dachs (red).
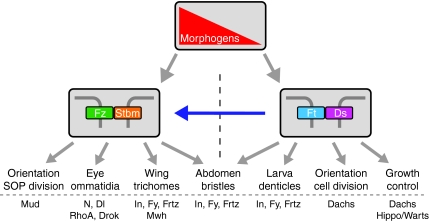
Relationships between the core and Ft/Ds pathways in Drosophila. Both the core and Ft/Ds pathways can respond independently to upstream morphogenetic patterning information (gradient in red). The core pathway (represented by heterophilic Fz-Stbm binding) interacts directly with effectors to control polarity of trichomes in the wing, the orientation of sensory organ precursor (SOP) divisions, and the DV polarity and rotation of ommatidia in the eye. In these contexts the Ft/Ds pathway can alter polarity via the core pathway (blue arrow) by an unknown mechanism. The Ft/Ds pathway can act independently of the core pathway to mediate effects on growth control, cell division orientation in the wing and eye discs, and the polarity of larval denticles. The Ft/Ds pathway acts in parallel with the core pathway to control trichome and bristle orientation in the abdomen. Example effectors mediating these effects are shown at the bottom. Dl, Delta; Drok, Rho kinase; Ds, Dachsous; Frtz, Fritz; Ft, Fat; Fy, Fuzzy; Fz, Frizzled; In, Inturned; Mud, Mushroom Body Defective; Mwh, Multiple Wings Hairs; N, Notch; Stbm, Strabismus.
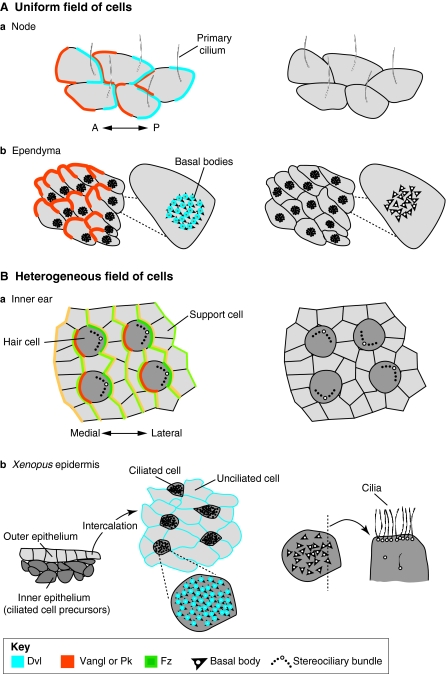
Organisation of vertebrate planar polarity in uniform and heterogeneous fields of cells. (A) Planar polarity in uniform fields of cells. In all panels, protein distributions are schematised on the left where known: Dvl (Dishevelled; blue), Vangl (Vang-like) or Pk (Prickle; orange), Fz (Frizzled; green). Surface views shown except where noted. The arrangements of polarised cells as seen in planar polarity mutants are illustrated on the right. In (a) the vertebrate node and (b) the ependymal lining of the brain, identically polarised cells point in one direction, as revealed by the asymmetric position of primary cilia in the node and patches of cilia in the ependyma (translational polarity). Planar polarity also exists at the subcellular level – i.e. rotational polarity – as revealed by the alignment of basal bodies (arrowheads) in individual ependymal cells (b). Cilia remain in the centre and/or fail to align with each other when planar polarity is not properly established. (B) In heterogeneous fields of cells, such as (a) the mouse inner ear and (b) Xenopus epidermis, polarised cells interdigitate with cells that show no outward polarisation. In the inner ear, core proteins are asymmetrically distributed in both hair cells (dark grey) and support cells (light grey); protein distributions are represented in lighter shades in support cells. In polarity mutants, hair cells are not uniformly oriented. In the epidermis, ciliated cells are born in the inner epithelium and then intercalate with unciliated cells in the outer epithelium (shown in cross-section), creating a mosaic of polarised and unpolarised cells in the mature epithelium (shown in surface view). Dvl and Vangl proteins may localise to basal bodies, which normally all point in the same direction. In planar polarity mutants, ciliary orientation is disrupted; defects in ciliogenesis and the apical localisation of the basal bodies have also been reported, as illustrated for one cell shown in cross-section on the right. A, anterior; P, posterior.
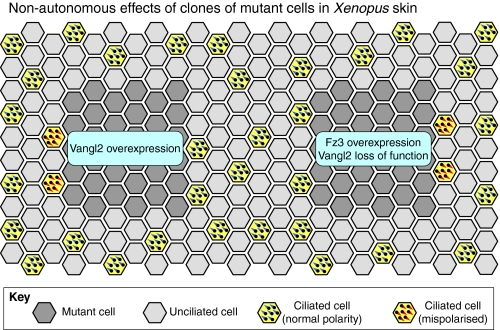
Non-autonomous planar polarity phenotypes in the Xenopus epidermis. Grafting experiments in the Xenopus epidermis reveal that core proteins mediate non-autonomous cell-cell interactions that control the polarity of neighbouring tissue. Grafts of cells with altered core protein expression are indicated in dark grey. Surrounding cells are either multiciliated (yellow) or non-ciliated (light grey). Multiciliated cells are normally polarised to direct fluid flow posteriorly and ventrally (bottom left of the diagram, basal bodies shown in blue). However, in wild-type cells on the anterior edge of grafts that overexpress Vang-like2 (Vangl2), or on the posterior edge of grafts that overexpress Frizzled3 (Fz3) or that have reduced Vangl2 activity, cells produce cilia with inverted polarity (basal bodies shown in red). Anterior is towards the left.
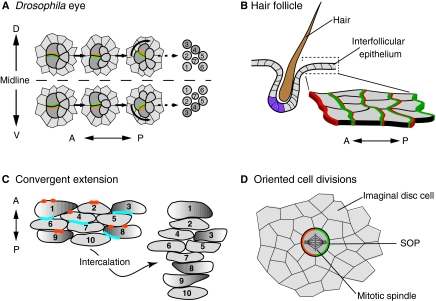
Planar polarity organisation in multicellular structures and during morphogenesis. Planar polar organisation is also evident in multicellular structures, such as in (A) Drosophila ommatidia and (B) mammalian hair follicles. (A) In the developing fly eye, planar polarisation of core proteins (green and orange) determines which cells will be R3 and R4 photoreceptors (dark grey) and the direction of ommatidium rotation [which at this stage consists of the R3 and R4 photoreceptors, plus R7, R2 and R5 (light grey)]. (Right) The final adult orientation of R1-R7. (B) In mammalian skin, hair follicles comprise multiple cell types that differ in gene expression (purple) and morphology along the anteroposterior (AP) axis. Core proteins (green and orange) are asymmetrically localised across the epithelium, including in the interfollicular cells (as shown in the magnified view). Planar polarity also directs cell movements and oriented cell divisions that sculpt tissues during morphogenesis, as seen during (C) convergent extension in vertebrates and (D) sensory organ precursor (SOPs, dark grey) division in fly imaginal discs. (C) During convergent extension, cells extend along the mediolateral axis (illustrated by graded shading of a subset of cells) and intercalate with each other, causing elongation along the AP axis. Each cell is numbered before and after intercalation to highlight the changing cell-cell interactions. Dvl (Dishevelled) proteins (blue) are localised posteriorly, and Pk (Prickle; orange) anteriorly, although in some contexts, asymmetric localisation has been difficult to detect. (D) Asymmetric localisation of core proteins (green and orange) in SOPs determines the orientation of the mitotic spindle and hence cell division.
References
-
- Adler P. N. (1992). The genetic control of tissue polarity in Drosophila. BioEssays 14, 735-741 - PubMed
-
- Adler P. N. (2002). Planar signalling and morphogenesis in Drosophila. Dev. Cell 2, 525-535 - PubMed
-
- Adler P. N., Krasnow R. E., Liu J. (1997). Tissue polarity points from cells that have higher Frizzled levels towards cells that have lower Frizzled levels. Curr. Biol. 7, 940-949 - PubMed
-
- Adler P., Charlton J., Liu J. (1998). Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype by altering frizzled signaling. Development 125, 959-968 - PubMed
-
- Adler P. N., Liu J., Charlton J. (2000). Cell size and the morphogenesis of wing hairs in Drosophila. Genesis 28, 82-91 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
