

Selective activation of the transcription factor ATF6 mediates endoplasmic reticulum proliferation triggered by a membrane protein
- PMID: 21521793
- PMCID: PMC3093499
- DOI: 10.1073/pnas.1101379108
Selective activation of the transcription factor ATF6 mediates endoplasmic reticulum proliferation triggered by a membrane protein
Abstract
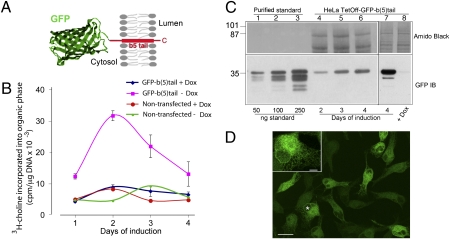
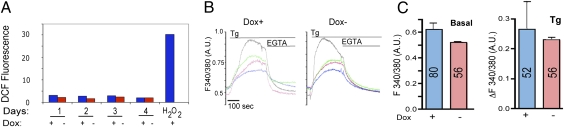
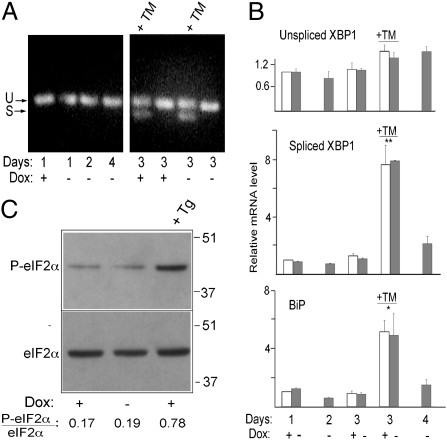
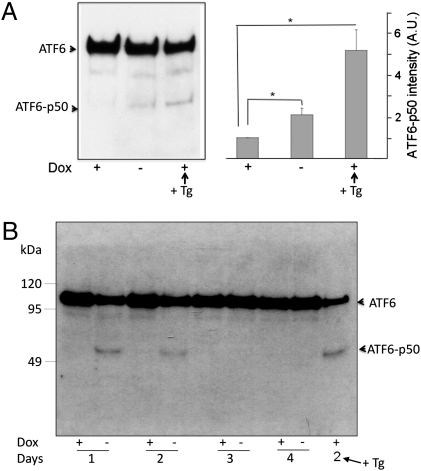
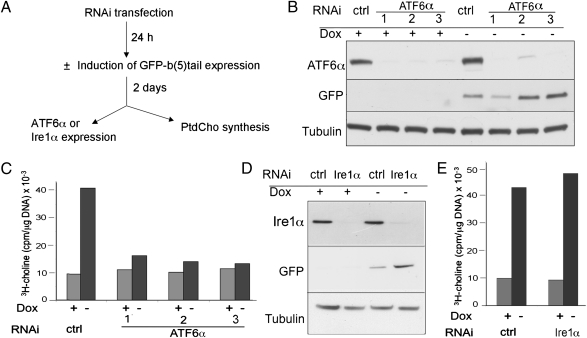
It is well known that the endoplasmic reticulum (ER) is capable of expanding its surface area in response both to cargo load and to increased expression of resident membrane proteins. Although the response to increased cargo load, known as the unfolded protein response (UPR), is well characterized, the mechanism of the response to membrane protein load has been unclear. As a model system to investigate this phenomenon, we have used a HeLa-TetOff cell line inducibly expressing a tail-anchored construct consisting of an N-terminal cytosolic GFP moiety anchored to the ER membrane by the tail of cytochrome b5 [GFP-b(5)tail]. After removal of doxycycline, GFP-b(5)tail is expressed at moderate levels (1-2% of total ER protein) that, nevertheless, induce ER proliferation, as assessed both by EM and by a three- to fourfold increase in phosphatidylcholine synthesis. We investigated possible participation of each of the three arms of the UPR and found that only the activating transcription factor 6 (ATF6) arm was selectively activated after induction of GFP-b(5)tail expression; peak ATF6α activation preceded the increase in phosphatidylcholine synthesis. Surprisingly, up-regulation of known ATF6 target genes was not observed under these conditions. Silencing of ATF6α abolished the ER proliferation response, whereas knockdown of Ire1 was without effect. Because GFP-b(5)tail lacks a luminal domain, the response we observe is unlikely to originate from the ER lumen. Instead, we propose that a sensing mechanism operates within the lipid bilayer to trigger the selective activation of ATF6.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol. 2007;8:519–529. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
