

Ambivalent role of the innate immune response in rabies virus pathogenesis
- PMID: 21525357
- PMCID: PMC3126514
- DOI: 10.1128/JVI.00302-11
Ambivalent role of the innate immune response in rabies virus pathogenesis
Abstract
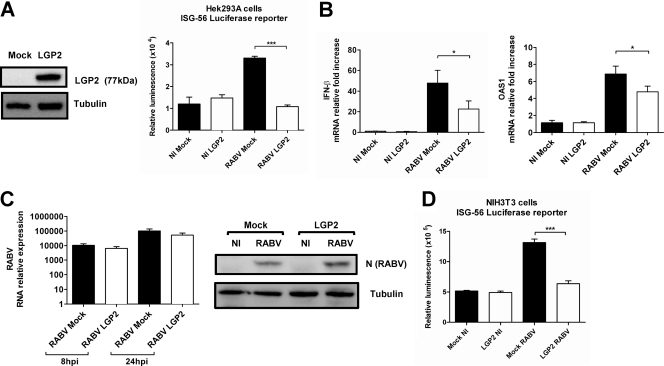
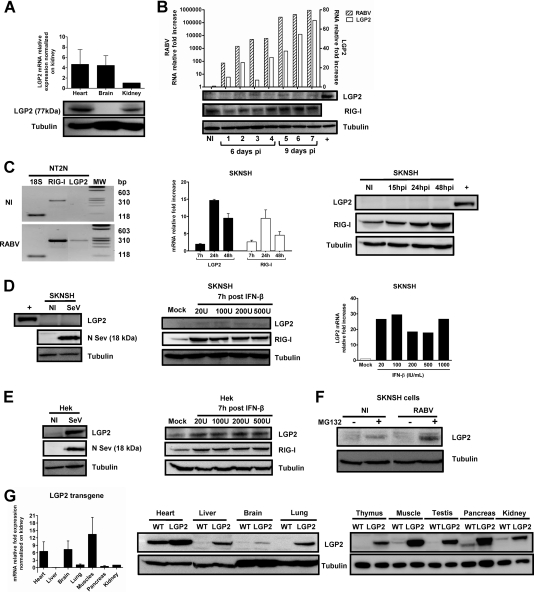
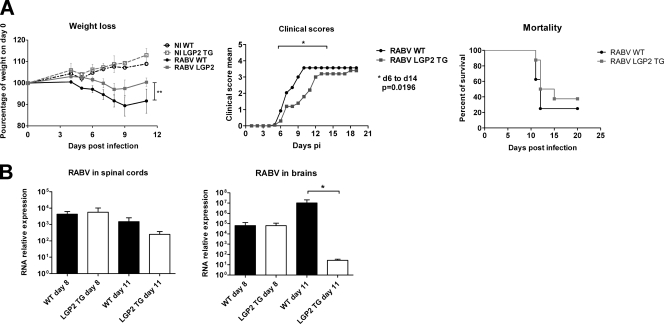
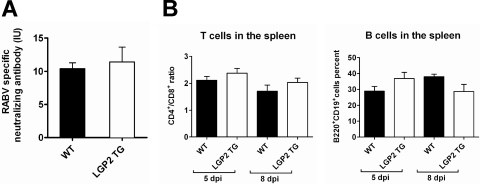
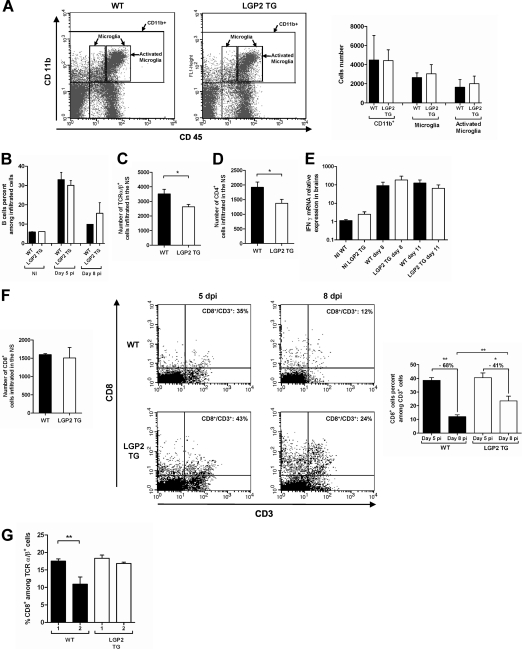
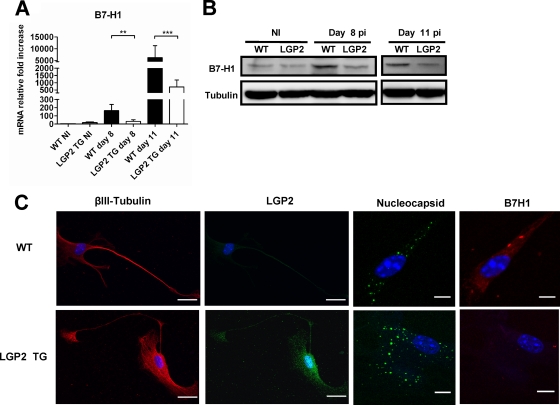
The neurotropic rabies virus (RABV) has developed several evasive strategies, including immunoevasion, to successfully infect the nervous system (NS) and trigger a fatal encephalomyelitis. Here we show that expression of LGP2, a protein known as either a positive or negative regulator of the RIG-I-mediated innate immune response, is restricted in the NS. We used a new transgenic mouse model (LGP2 TG) overexpressing LGP2 to impair the innate immune response to RABV and thus revealed the role of the RIG-I-mediated innate immune response in RABV pathogenesis. After infection, LGP2 TG mice exhibited reduced expression of inflammatory/chemoattractive molecules, beta interferon (IFN-β), and IFN-stimulated genes in their NS compared to wild-type (WT) mice, demonstrating the inhibitory function of LGP2 in the innate immune response to RABV. Surprisingly, LGP2 TG mice showed more viral clearance in the brain and lower morbidity than WT mice, indicating that the host innate immune response, paradoxically, favors RABV neuroinvasiveness and morbidity. LGP2 TG mice exhibited similar neutralizing antibodies and microglia activation to those of WT mice but showed a reduction of infiltrating CD4(+) T cells and less disappearance of infiltrating CD8(+) T cells. This occurred concomitantly with reduced neural expression of the IFN-inducible protein B7-H1, an immunoevasive protein involved in the elimination of infiltrated CD8(+) T cells. Our study shows that the host innate immune response favors the infiltration of T cells and, at the same time, promotes CD8(+) T cell elimination. Thus, to a certain extent, RABV exploits the innate immune response to develop its immunoevasive strategy.
Figures


References
-
- Baloul L., Camelo S., Lafon M. 2004. Up-regulation of Fas ligand (FasL) in the central nervous system: a mechanism of immune evasion by rabies virus. J. Neurovirol. 10:372–382 - PubMed
-
- Blondel D., et al. 2002. Rabies virus P and small P products interact directly with PML and reorganize PML nuclear bodies. Oncogene 21:7957–7970 - PubMed
-
- Brehin A.-C., et al. 2008. Dynamics of immune cell recruitment during West Nile encephalitis and identification of a new CD19+ B220− BST-2+ leukocyte population. J. Immunol. 180:6760–6767 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
